compcogneuro/book: パート II: 脳領域
このページは外部資料の日本語訳です。原文の見出し順と本文順を保ち、コード・URL・出典表記はできるだけ原形のまま残しています。
出典とライセンス
原典: https://github.com/compcogneuro/book/blob/main/chapter-05.md
ライセンス: CC BY 4.0。このページは日本語翻訳であり、変更点は翻訳とサイト内整形です。
bibfile: ccnlab.bib —
パート II: 脳領域
本書の第 1 部では、個々のニューロンの活性化ダイナミクスからニューロンのネットワーク、そして自己組織化とエラー駆動の両方の方法でニューロンを構成する学習メカニズムまで、基本的なニューロン メカニズムのツールキットを開発しました。第 II 部の開始では、広範囲の認知現象の探索への移行を開始します。この道に沿った重要な基礎的なステップとして、この章では、文献で一般的に合意されている内容にほぼ対応する、比較的議論の余地のない方法で、脳の機能組織全体の全体像を提供することを試みます。これは、脳のさまざまな領域がどのように連携してさまざまな認知機能を実行するかを広範なレベルで理解し、後続の章でより具体的なモデルをより大きな全体的なフレームワークに位置づけるのに役立ちます。
私たちは後続の章と同じ順序で進み、基本的な知覚システムから始めて、さまざまな形の学習と記憶 (エピソード記憶における海馬の役割を含む) の探求に進みます。次に、基本的な運動システムに目を向け、それらに基づいて実行機能を調べます。最後に、言語を調査する際に、これらすべての認知システムの機能を構築および拡張します。
いつものように、生物学の基本的な基礎である脳の肉眼的な解剖学から始めます。
脳の機能解剖学をナビゲートする
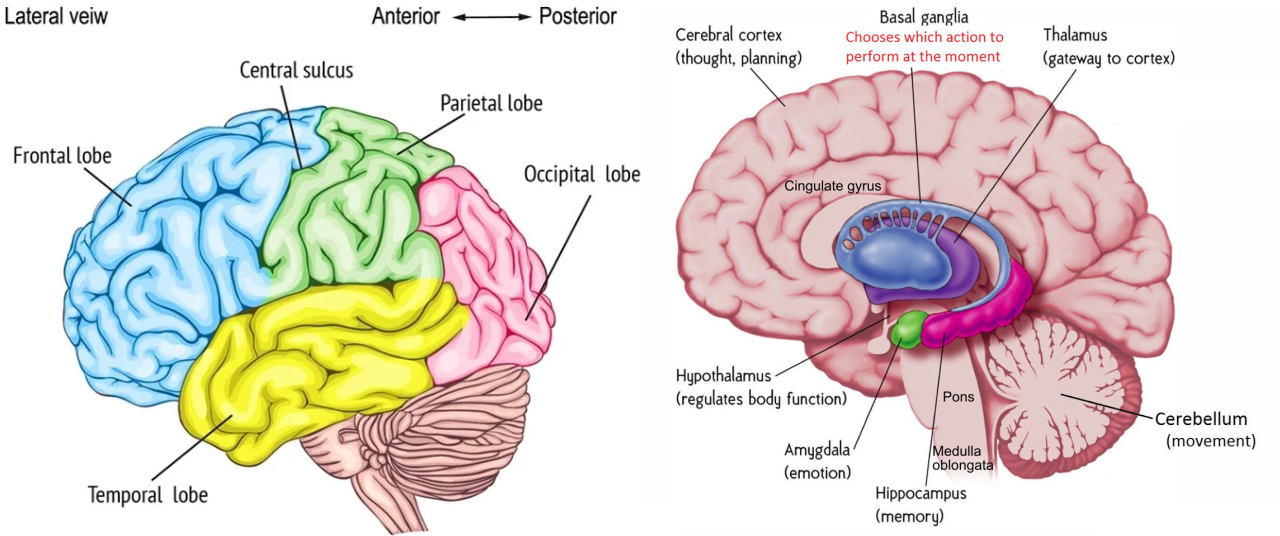
[@fig:fig-brain-anatomy] は、脳の「全体的な」(実際には非常に美しく、驚くべき!) 解剖学的構造を示しています。外側の部分は新皮質の「しわのあるシート」(その上に私たちの思考が置かれます)であり、主要な葉のすべてが示されています。これは、私たちの複雑な認知機能のほとんどが発生する場所であり、本文のこの時点で私たちが焦点を当ててきたことです。脳の残りの部分は新皮質の内側にあり、いくつかの重要な領域が図に示されています。これらは一般に 皮質下 脳領域と呼ばれており、次のようないくつかの領域が計算モデルに含まれています。
-
海馬 — この脳領域は、実際には「古皮質」と呼ばれる皮質の「古代」形態であり、出来事や事実に関する新しい「日常」の記憶 (エピソード 記憶と呼ばれる) を学習する際に海馬がどのように重要な役割を果たしているかを 記憶 の章で説明します。
-
扁桃体 — この脳領域は、感情的に顕著な刺激を認識し、それについて脳の他の部分に警告するために重要です。これについては 運動制御と強化学習 の章で詳しく説明します。そこでは、報酬 (および罰) に基づいて運動 (および認知) 行動を強化する上で重要な役割を果たします。
-
小脳 — この巨大な脳構造には脳内のニューロンの 1/2 が含まれており、運動調整において重要な役割を果たしています。また、ほとんどの認知タスクでも活動しますが、認知におけるその機能的役割が何であるかを正確に理解することは、ややとらえどころのないままです。これについては、モーター制御と強化学習 の章で説明します。
-
視床 — 新皮質に向かう感覚情報の主要な経路を提供し、注意、覚醒、その他の調節機能にも重要であると考えられます。 知覚と注意の章では視覚視床の役割を、運動制御と強化学習の章では運動視床の役割を探ります。
-
大脳基底核 — これは、運動制御と強化学習 の章、および 実行機能 の章で重要な役割を果たす皮質下領域の集合です。これは、皮質が「提案」する特定の行動を実行するかどうか、また前頭前野の認知計画を更新するかどうかについて、最終的な「Go」の判断を下すのに役立ちます。これらの選択を行うためのポリシーは、以前の強化/懲罰の歴史に基づいて学習されます。
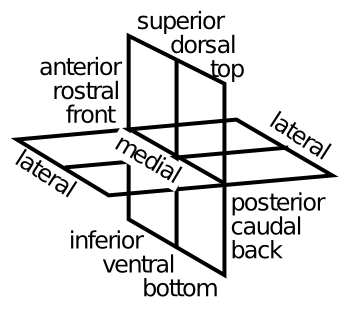
[@fig:fig-location-terms] は、解剖学者が脳のさまざまな部分について話すために使用する用語を示しています。これらの用語に慣れておくことは良いことです。今すぐ活用してみましょう。
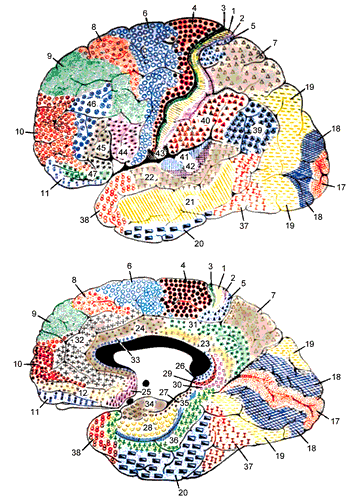
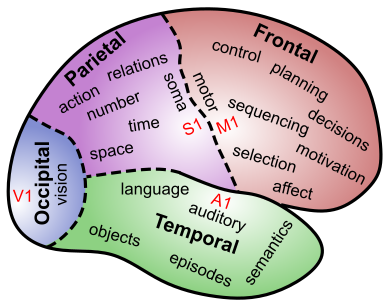
[@fig:fig-brodmann-areas-color; @fig:fig-brodmann-areas-3d-color] は、ブロードマン領域 の観点から、新皮質の構造の詳細を示しています。これらの領域は、解剖学的差異 (主に、ネットワーク 章で説明した異なる皮質層の厚さの違い) に基づいて、コルビニアン ブロードマンによって特定されました。このレベルの詳細についてはあまり言及しませんが、これらの数値のいくつかを学習することは、認知神経科学の主要な文献を読めるようになるために良いアイデアです。ここでは、皮質葉の機能の簡単な概要を示します ([@fig:fig-cortex-lobes])。
-
後頭葉 — これには、新皮質の最後部に位置する 一次視覚野 (V1) (ブロードマン野 17 または BA17) と、そこから放射状 (前方) に広がる高次の視覚野が含まれます。明らかに、その主な機能は視覚処理です。
-
側頭葉 — 後頭葉から出発し、視覚処理の経路は下側頭葉(IT)に下り、そこで視覚物体が認識されます。一方、上側頭皮質には **一次聴覚野 (A1) と、関連する高次の聴覚領域および 言語処理 領域が含まれています。したがって、側頭葉(両側に 1 つ)は、物体の視覚的外観が言語ラベルに変換される場所(またはその逆)であり、また、私たちが読み方を学ぶ場所でもあります。側頭葉の最も前方の領域は、意味的知識、つまり弁護士や政府などの高度な理解や、学校で学ぶすべての優れた知識にとって重要であるようです。 内側側頭葉 (MTL) 領域は海馬に移行しており、ここの領域は人生の出来事の記憶 (エピソード記憶) の保存と検索においてますます重要な役割を果たしています。より深い意味学習を行わずに丸暗記をしているとき、MTL と海馬は懸命に働きます。最終的に、物事をより深く体系的に学習すると、それらは前側頭葉皮質 (および他の脳領域も) にエンコードされます。要約すると、側頭葉には、事実、出来事、名前、顔、物体、言葉など、私たちが意識的に認識している膨大な量の情報が含まれています。大まかな特徴の 1 つは、側頭葉が無数の方法で世界を分類するのが得意であるということです。
-
頭頂葉 — 側頭葉とは対照的に、頭頂葉ははるかに曖昧で潜在意識にあります。これは、空間的位置 (つまり、IT のどの経路を補完する どこ 経路) をエンコードするのに重要であり、その特定の部分が損傷すると、半空間無視という現象が生じます。人々は空間の半分全体を忘れてしまいます。しかし、その機能は単なる空間的な位置をはるかに超えています。これは、数値、数学、抽象的な関係、その他多くの「賢い」ものをエンコードするために重要です。より現実的なレベルでは、頭頂皮質は視覚情報が運動行動を導く主要な経路を提供し、方法経路として特徴付けられます。また、一次体性感覚皮質 (S1) も含まれており、運動動作を導き、情報を与えるためにも重要です。頭頂葉皮質の一部の部分では、ニューロンが異なる基準フレームの間で変換する役割を果たしており、たとえば、(体性感覚からの)身体上の空間的位置を視覚座標に変換します。また、視覚情報は、網膜上の活動のパターン (網膜局所座標)、または頭、体、または環境ベースの参照フレームの観点からエンコードできます。頭頂葉皮質の大まかな特徴の 1 つは、計量 情報、つまり側頭葉の離散的でカテゴリ的な性質とは対照的に、連続体に沿って変化する情報の処理に特化していることです。同様の区別は、脳の左側と右側の観点からよく議論されていますが、側頭部と頭頂部の観点から見た場合の証拠は、全体としてより強力です。
-
前頭葉 — これは一次運動野(M1)の後端から始まり、さらに進むと、M1および補足運動野(SMA)における低レベルの運動制御から、運動前野(高次運動野)にコード化されたより高次の行動シーケンスおよび偶発的行動に至るまで、より高次の運動制御の階層が存在する。その先には、脳の執行機関として知られる前頭前皮質(PFC)があります。ここがすべてのハイレベルなショットが呼び出される場所であり、ここで大きな計画が整理され、基本的な動機や感情の影響を受けて、実際に次に何をすべきかを決定します。 PFC には後部と前部の組織もあり、より前方の領域がより高いレベルの長期的な計画と目標を符号化しています。 PFC の最も前方の領域 (前頭極) は、最も抽象的で難しい形式の認知的推論にとって、特に重要であるようです。パズルを解こうと懸命に努力しているときや、GRE や IQ テストの難しい問題を解決しようとしているときです。前頭皮質の内側領域と腹側領域は、*感情と動機付けにとって特に重要です。たとえば、眼窩前頭皮質 (OFC)** は、与えられた刺激や起こり得る結果がどれほど報われるかについての情報を維持および操作するために重要であるようです (この情報を学習し表現するのを助けるために、扁桃体から強力な入力を受けます)。 前帯状皮質 (ACC) は、現在の状態での将来の行動に関連する困難、不確実性、または失敗の可能性など、行動の結果をコード化するのに重要です (スキー場で実行されている二重の黒いダイヤモンドを見下ろすと点灯します!)。 OFC と ACC は両方とも、他の前頭運動計画領域との相互作用を介して、また大脳基底核との相互作用を介して選択に影響を与えることができます。 腹内側 PFC (VMPFC) は、多くの皮質下領域と相互作用して、心拍数、呼吸、神経調節領域などの基本的な身体機能を制御し、その後、より広範囲に脳に影響を及ぼします (例: 腹側被蓋野 (VTA) や青斑核 (LC) は、ドーパミン と ノルアドレナリン を放出します。どちらも皮質全体に広範囲に影響しますが、特に前頭部に影響します。皮質)。前頭葉に関する最大の謎は、「幹部」などの用語を使わずに、前頭葉がこれらすべての驚くべきことをどのように行うかをどうやって理解するかということです。なぜなら、ピンストライプのスーツを着た小さな男がそこに座っているはずがないからです。それはすべて単なるニューロンです!
主要な脳領域の比較と対比
| 学習信号 | ダイナミクス | |||||
|---|---|---|---|---|---|---|
| エリア | 報酬 | エラー | 自己組織 | 区切り文字 | インテグレーター | アトラクター |
| 大脳基底核 | +++ | — | — | ++ | - | — |
| 小脳 | — | +++ | — | +++ | — | — |
| 海馬 | + | + | +++ | +++ | — | +++ |
| 新皮質 | ++ | +++ | ++ | — | +++ | +++ |
表: 脳の 4 つの主要領域にわたる学習メカニズムと活動/表現のダイナミクスの比較。 +++ は、そのエリアに確実に特定の特性があることを意味し、+ が少ない場合は、この地物に対する信頼性および/または重要性が低いことを示します。 --- は、そのエリアに特定のプロパティが明らかに存在しないことを意味し、やはり - が少ない場合は、信頼度または重要性が低いことを示します。
[@tbl:table-learning] は、採用される学習ルールと活性化ダイナミクスに基づいた 4 つの主要な脳領域の比較を示しています。報酬主導型学習についてはまだ説明していません。 運動制御と強化学習 の章でそれを行いますが、今のところ、どの活性化パターンが報酬信号 (ドーパミン レベルなど) を最大化するかを予測する学習として考えてください。
大脳基底核、小脳、海馬の進化的に古い領域は、別々の形式の活性化ダイナミクスを採用しています。これは、たとえ多少似た入力であっても、構造内の神経活動のより別々のパターンにマッピングする傾向があることを意味します。これは、特定の入力に対する特定の答えを暗記することに似た、非常に保守的で堅牢な戦略です。効率的ではなく、新しい状況にうまく一般化できないとしても、問題なく機能する可能性が高いです。これらの構造はそれぞれ、この全体的な分離のダイナミックさの中で、さまざまな形の学習を最適化していると見ることができます。大脳基底核は、報酬の期待と結果に基づいた学習に特化しています。小脳は、シンプルかつ効果的な形式のエラー駆動型学習 (基本的には 学習 の章で説明するデルタ ルール) を使用します。そして、海馬はヘビアンスタイルの自己組織化学習にさらに依存しています。したがって、海馬は、エラーや報酬に関係なく、常に新しいエピソード記憶をエンコードしています(ただし、表の弱い+記号で示されているように、これらは確かに学習速度を調整する可能性があります)。一方、大脳基底核は、潜在的な報酬またはその欠如に基づいて運動行動を選択することを学習しています(また、行動選択のタイミングを調節するための制御システムでもあります)。一方、小脳は、相対的な感覚フィードバックの違いから生成されるエラー信号を使用して、それらの運動行動を迅速に実行することを学習しています。モータープランへ。これら 3 つのシステムを総合すると、生物が生き残り、環境に適応するための基本的なニーズを少なくともある程度は満たすのに十分です。
海馬には、大脳基底核や小脳に存在するものを超える重要な革新が 1 つ導入されています。それは、アトラクターダイナミクスを備えていることです。特に、CA3 ニューロン間の反復接続は、ネットワーク 章で説明したように、パターン補完を介して、以前にエンコードされた記憶を取得するために重要です。この技術革新の代償として、てんかんの活動のダイナミクスを防ぐために、興奮と抑制のバランスを正確に維持する必要があります。実際、海馬は、少なくとも人々において、てんかん活動の最も一般的な発生源です。
進化的に古いシステムを背景として、新皮質はいくつかの重要な革新を表しています。活性化ダイナミクスの観点から見ると、これは海馬からのアトラクターのダイナミックなイノベーションを基礎としており(海馬が古代の「原」皮質を表すことを考えると、当然のことです)、これに加えて、すべてを常に分離したままにするのではなく、経験全体を統合して一般性を抽出する表現を開発する強力な能力が追加されます。この統合機能の代償として、システムが間違った種類の一般化を形成できるようになり、全体的な動作が悪化する可能性があります。しかし、以前の学習を新たな状況に適用する強力な能力をシステムに与えることで、明らかに利点がリスクを上回ります。学習メカニズムに関しては、新皮質は 3 つの主要な学習形式をすべてしっかりとブレンドし、利用可能なすべての学習信号の最良のものを 1 つのシステムに統合します。
第 II 部の残りの各章では、さまざまな認知機能に焦点を当てます。以下に各章のプレビューを示します。
知覚と注意: 何がどこなのか
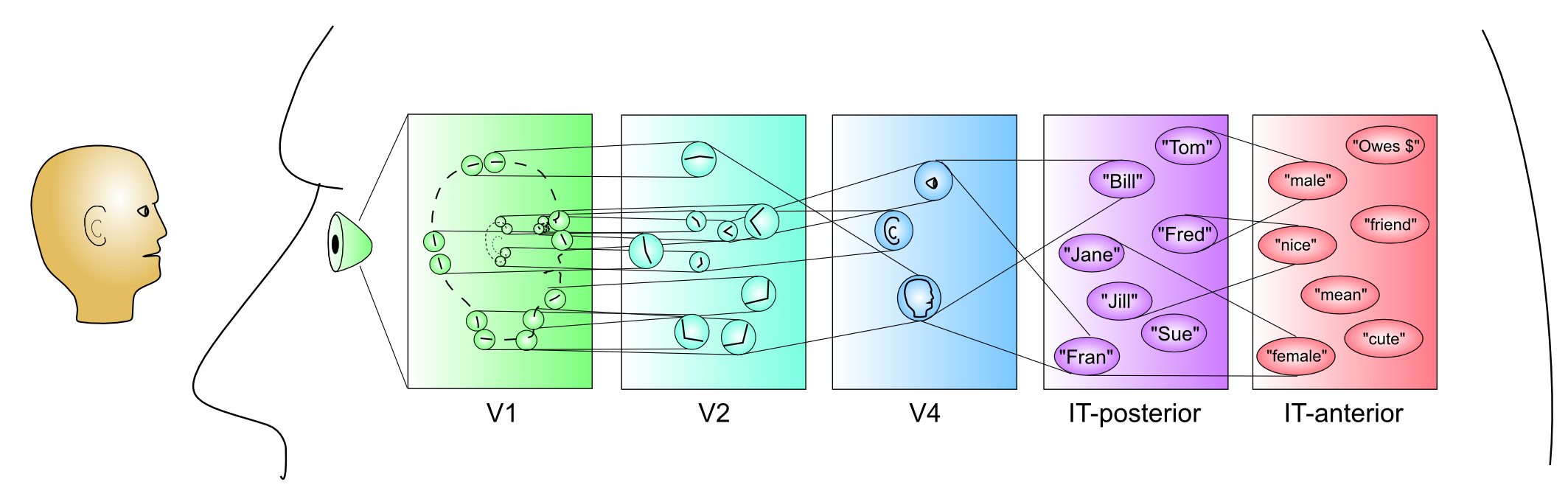
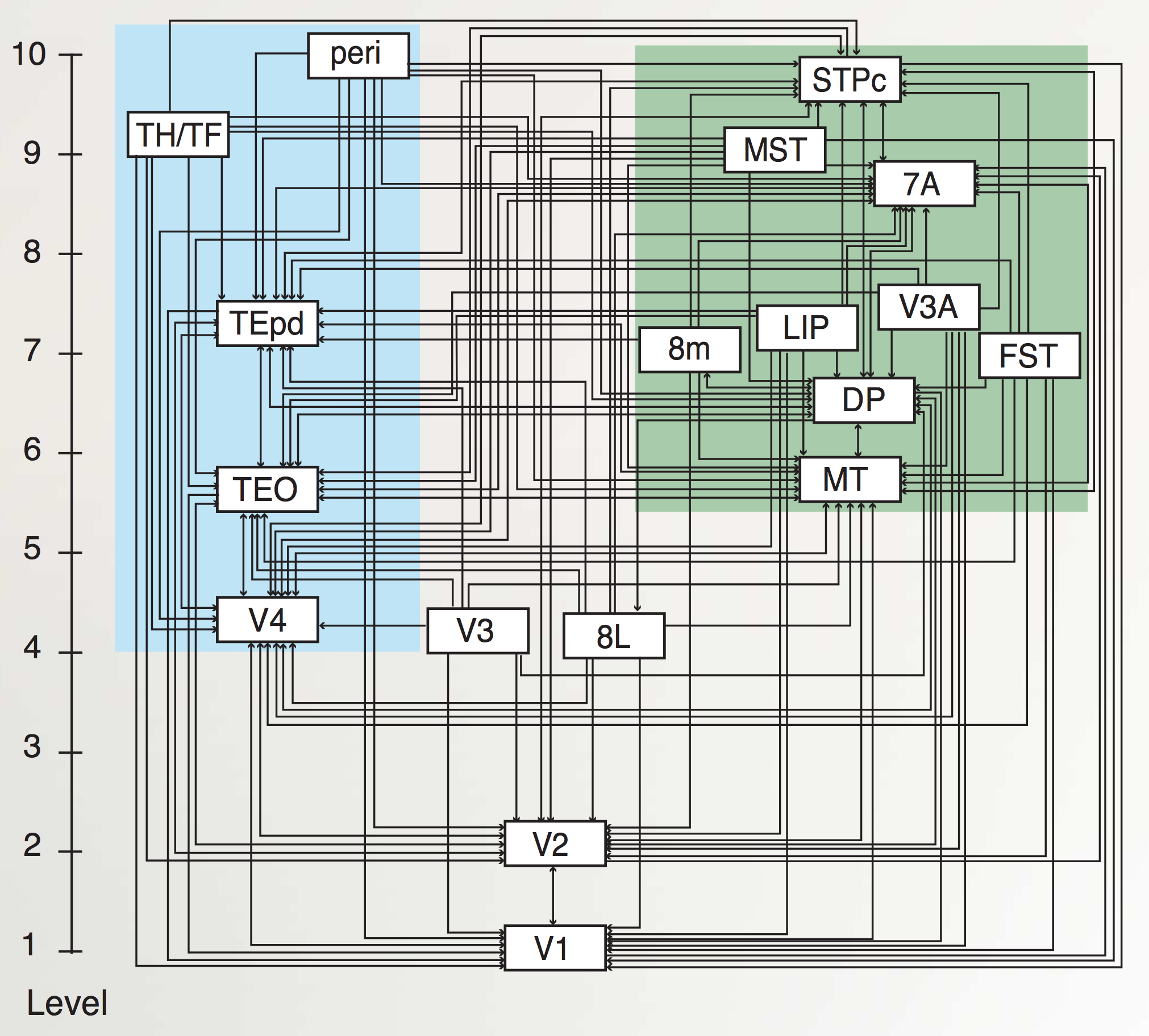
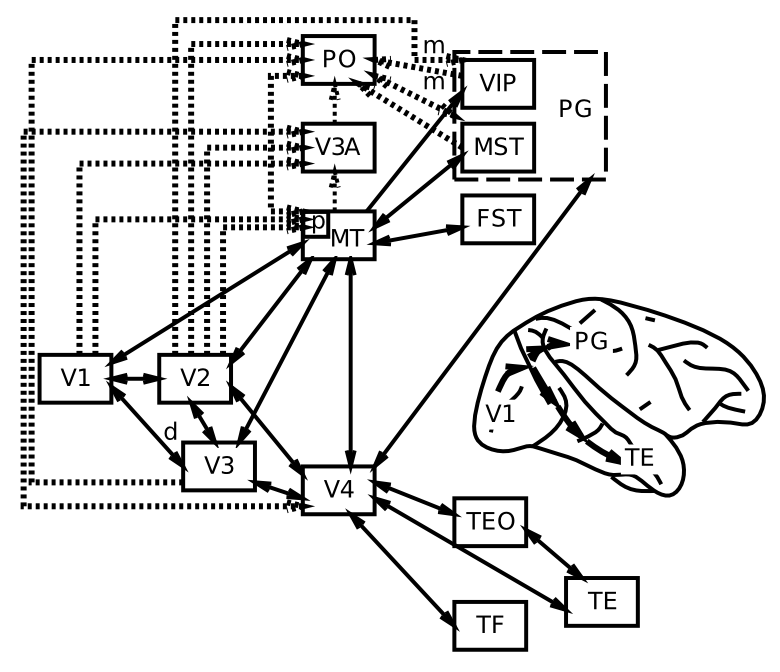
ネットワーク の章で説明したように、知覚システムは、階層的に組織された神経検出器の層の力を示す優れた例を提供します。 [@fig:fig-category_hierarch_dist_reps-5] はこのプロセスを要約しており、処理の各段階の下に関連する皮質領域が示されています。 [@fig:fig-markov_cort_hier-5] は、すべての主要な視覚野 [@MarkovVezoliChameauEtAl14; @FellemanVanEssen91] の実際の解剖学的接続パターンの現在の最良の推定値を示しており、情報が実際に脳内で階層的に処理されていることを示しています (ただし、厳密な階層の外側にも多くの相互接続があります)。 [@fig:fig-vis_system_bio-5] は、これらの領域を解剖学的位置に配置し、視覚処理 [@UngerleiderMishkin82] における 何を対どこ (腹側と背側) の分割をより明確に示します。以下は、視覚経路の何側を上る情報の流れの簡単な概要です([@fig:fig-markov_cort_hier-5]の左側に図示):
-
V1 — 一次視覚野。さまざまな配向角度に沿ったエッジ (照明の遷移) に応答する配向エッジ検出器の観点から画像をエンコードします。 知覚と注意 の章では、これらのエッジ検出器が、自然画像の信頼できる統計によって駆動される自己組織化学習を通じてどのように開発されるかを見ていきます。
-
V2 — 二次視覚野。より複雑な形状を検出するための基盤を提供する、他の多くの基本的な視覚機能 (例: 3D 深さの選択性、基本的なテクスチャなど) とともに、交差点や接合部のボキャブラリーを開発するためのエッジ検出器の組み合わせをエンコードします。これらの V2 ニューロンは、これらの特徴をより広範囲の場所でエンコードし、最終的に IT ニューロンが視野内のどこに現れるかに関係なく、物体を認識できるようになるプロセスを開始します (つまり、不変 物体認識)。
-
V4 — より広い範囲の位置 (およびサイズ、角度など) にわたって、より複雑な形状特徴を検出します。
-
IT-posterior (PIT) — 広範囲の位置、サイズ、角度にわたってオブジェクト全体の形状を検出します。たとえば、側頭葉の下面の紡錘状回の近くには、紡錘状顔面領域 (FFA) と呼ばれる領域があり、特に顔に反応すると思われます。ただし、ネットワーク の章で見たように、オブジェクトは IT の幅広い領域にわたって分散表現でエンコードされます。
-
IT-anterior (AIT) — ここでは、視覚情報が本質的に非常に抽象的かつ 意味論的になります — 図に示すように、さまざまな人々、場所、物に関するあらゆる種類の重要な情報をエンコードできます。
知覚と注意 の章では、不変オブジェクト認識のモデルを検討し、この深い階層の検出器が学習を通じてどのように発達するかを示します。 言語 章は、このオブジェクト認識プロセスに基づいて、単語がどのように認識され、読書中に関連する言語運動出力に変換され、意味知識にも関連付けられるかを理解します。
視覚処理の 場所 は、中側頭葉 (MT)、腹側頭頂内皮質 (VIP)、外側頭頂葉内皮質 (LIP)、内側上側頭葉 (MST) などの領域を含む頭頂皮質で発生します。これらの領域は、動き、深さ、その他の空間的特徴の処理に重要です。上で述べたように、これらの領域は視覚入力を適切な運動出力に変換するためにも重要であり、Goodale と Milner はこれを how 経路 [@GoodaleMilner92] として特徴付けています。 知覚と注意 の章では、視覚的注意の文脈において、この背側経路が腹側経路とどのように相互作用し、たとえば半空間無視などの頭頂部損傷の特徴的な影響を生み出すのかを見ていきます。 また、見る (外側頭頂内皮質 (LIP) / 前頭眼野 (FEF))、 * 到達する * (VIP / 補足運動野 (SMA))、および * ナビゲートする * (後帯状皮質 (PCC) および後脾皮質 (RSC) を含む内側頭頂葉ネットワーク) [@KravitzSaleemBakerEtAl11; @RanganathRitchey12] に対応する 3 つの異なる頭頂葉経路についての証拠も増えています。
記憶: 側頭皮質と海馬
記憶というと、おそらく「昨日の夕食は何を食べましたか?」などが思い出されるでしょう。 「どうすれば人の名前をもっとよく覚えられるでしょうか?」思い当たる傾向があります。ただし、これらは記憶の 1 つのカテゴリにすぎません。実際、メモリはニューラル ネットワークのいたるところに存在します。すべてのシナプスにはメモリを保存する容量があり、特定の「メモリ」を暗号化して取得するには、何百万ものそのようなシナプスの協調動作が必要です。記憶には多くの分類がありますが、実際に知っておく必要がある唯一の分類は、ここで提供されている脳の機能組織と同じです。記憶は脳のあらゆる領域に埋め込まれており、その記憶の性質はその領域の動作と密接に関係しています。運動皮質は運動記憶を学習します。頭頂皮質は、運動能力、つまり野球の打ち方などのことを学習します(ヒント: ボールから目を離さないでください。頭頂皮質は視覚的な入力を必要とします!)。
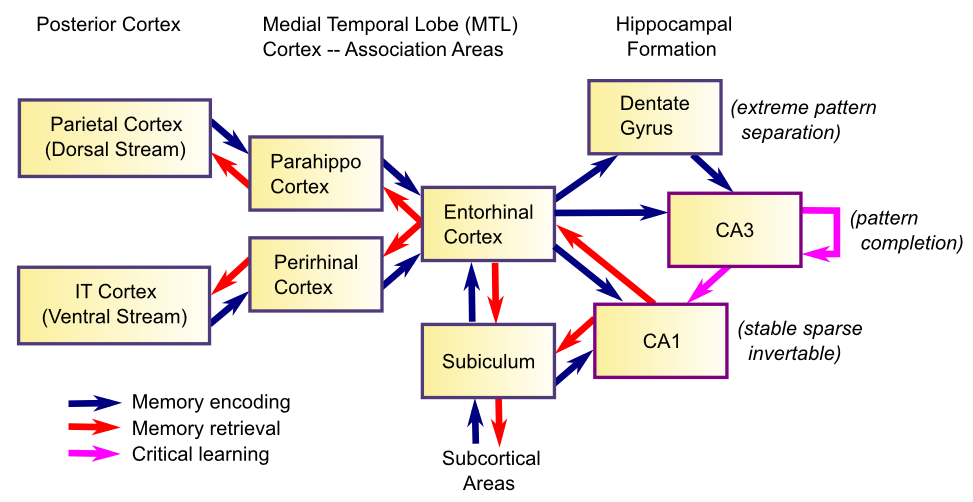
ただし、記憶の領域で非常に大きく迫っている脳の領域が 1 つあるため、そこに焦点を当ててみましょう。これは海馬で、以前に学習した情報[@fig:fig-hippo_mem_formation-5]にあまり干渉せずに、新しい情報を迅速に学習することに特に優れているようです。最近会った人の名前を思い出す必要があるときは、この海馬の急速な学習能力に頼ることになります。海馬系の神経特性が、この急速な学習能力を生み出すのに理想的に適していることがわかります。重要な神経特性の 1 つは、非常にスパースな表現を使用することです。これにより、パターン分離と呼ばれる現象が生じます。この現象では、ある記憶に関連付けられた神経活動パターンが、他の同様の記憶 [@Marr71; @McClellandMcNaughtonOReilly95] に関連付けられた神経活動パターンと大きく異なります。これにより、事前の学習への干渉が最小限に抑えられます。干渉は重複の関数として発生します。このパターン分離プロセスが、部分情報 [@OReillyMcClelland94] からの検索中に記憶を回復するためのパターン補完プロセスによってどのように補完されるかを見ていきます。
また、学習率が学習においてどのように重要な役割を果たすかについても見ていきます。明らかに、迅速に学習するには、速い学習速度が必要です。しかし、学習速度が遅い場合はどうなるのでしょうか?これにより、さまざまな経験を統合して、知恵 と 意味論的な知識 を生み出すことができることがわかりました。この学習速度の遅さは、大部分の新皮質の特徴です (これにより、大脳基底核は、最新の結果だけでなく、さまざまな経験にわたる各動作の肯定的な結果と否定的な結果の確率を学習することもできます) [@McClellandMcNaughtonOReilly95]。興味深いことに、学習速度が遅い場合でも、新皮質は、認識記憶 (つまり、明確なエピソード記憶なしで、何かを馴染みのあるものとして認識する能力) を促進する プライミング効果 や 慣れ親しんだシグナル の形で、1 回の学習試行で測定可能な効果を示すことができます。この形式の認識記憶は、鼻周皮質を含む内側側頭葉(MTL)領域に依存しているようです。ある試みの行動学習の別の形式には、アトラクター状態における記憶 (前頭前野の作業記憶) の能動的な維持をサポートするメカニズムが含まれます。この形式のメモリでは重みの変更はまったく必要ありませんが、インスタンス間の動作パフォーマンスに急速に影響を与える可能性があります。
運動制御: 頭頂葉、運動皮質、大脳基底核、小脳
頭頂方法経路を前方に運び、頭頂皮質を通って背側経路に沿って進む視覚情報は直接前頭皮質に向かい、そこで一次運動野の運動ニューロンを駆動し、筋肉を直接駆動して明白な運動動作を生み出すことができます。これにより、すべての行動の中核にある重要な感覚と運動のループが完成します。運動制御には、大脳基底核や小脳などの多くの皮質下の脳領域も大きく関与します。これらの領域の大まかな役割分担は次のとおりです。
-
新皮質 (頭頂葉から前頭葉) — 感覚情報の高レベルの計量処理を行い、複数のモダリティを統合し、必要に応じて異なる参照フレーム間で変換して、現在の感覚環境に対する可能な応答の範囲に到達します。
-
大脳基底核 — 感覚入力と前頭皮質で「考慮」されている潜在的な反応の両方を受け取り、脱抑制性の Go 信号をトリガーして、可能なアクションの ** 最良の** を可能にして閾値を超え、実際に行動 [@Mink96; @Frank05] を推進することができます。 行動選択のこのプロセスは強化学習によって形成されます。大脳基底核はドーパミンに満ちており、報酬と罰に応じて学習を促進し、選択プロセス自体の速度にも影響します[@SuttonBarto98; @MontagueDayanSejnowski96]。したがって、大脳基底核は、報酬をもたらす可能性が最も高く、罰をもたらす可能性が最も低い行動を選択します。 扁桃体は、報酬と罰に関連する感覚信号に反応してこれらのドーパミン信号を駆動する上で重要な役割を果たします。
-
小脳 — は頭頂皮質および運動皮質と深く相互接続されており、シンプルかつ強力な形式のエラー駆動型学習を使用して、感覚入力と運動出力の間の高解像度計量マップを取得することができます。したがって、感覚情報と運動フィードバック情報を適切に統合して効率的かつ制御された方法で動作することは、スムーズで調整された運動動作を生成するために重要です。また、学んだことを頭頂皮質と運動皮質に教える役割も果たしている可能性があります。
運動制御と強化学習 の章では、基本的な動作選択タスクにおいて、ドーパミン信号が大脳基底核の学習とパフォーマンスをどのように形成するかを見ていきます。次に、物体を固定するために目と頭の協調的な動きを実行する仮想ロボットにおける小脳の運動学習の興味深いモデルを探索します。このモデルは、小脳の学習に必要なエラー信号がどのように自然に発生するかを示しています。
興味深いことに、これらの「低レベル」の運動制御システムはすべて、最終的には「高レベル」の実行機能システム (前頭前野など) に組み込まれるため、運動制御を特に認知領域として考えていない人もいますが、実際には、最高レベルの認知機能の一部を理解するための強固な基盤となります。
実行機能: 前頭前皮質と大脳基底核
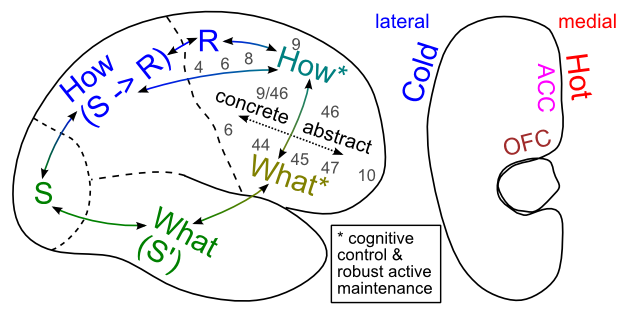
私たちは前頭葉皮質と大脳基底核の運動制御機能に基づいて、これら 2 つの領域がどのように相互作用して高度な実行機能をサポートするかを理解します。また、前頭前皮質の腹側領域と背側領域がどのように機能的に組織されているかを理解するために、後皮質の機能分割に基づいて構築します。 [@fig:fig-cortical_fun_org-5] は、これがどのように発生するかを示す全体的な図を示しています。また、外側表面が「コールド」認知機能とより関連しており、内側表面が「ホット」感情および動機付けの処理により深く関与していることも示しています。
PFC が後皮質の処理に対して トップダウンの認知制御をどのように提供できるかを見ていきます。典型的な例はストループ タスクです。
次に、PFC と BG がどのように相互作用して、システムが複数の情報を「頭の中に」保持し、既存の情報を維持し続けながら一部の情報を個別に更新できるようにする、動的にゲートされた作業記憶システムを生成する方法を検討します。このシステムにおけるBGの役割は、運動野ではなくPFCと非常によく似た回路で相互作用することにより、運動制御におけるBGのより確立された役割に基づいています。どちらの場合も、BG は、特定の前頭皮質の「アクション」を実行するかどうかを決定するためのゲート信号を提供します。ただ、PFC 動作は運動皮質よりも認知的であり、作業記憶状態や目標、計画などの更新などが含まれます。これらの PFC 表現は、更新されると、上で述べたトップダウンの認知制御を提供できるため、BG 運動回路での動作選択を形作ることができますが、感覚皮質の課題関連機能への注意にも影響を与えます。興味深いことに、どの認知行動を実行するかを強化するメカニズム(作業記憶を更新するかどうか、特定の機能に注意を向けるか、高レベルの計画を開始するかなど)は、運動制御の中心となる非常によく似たドーパミン作動性強化学習メカニズムに依存しているようです。このフレームワークは、モチベーションと行動の間の確立されたリンクと非常によく似た、モチベーションと認知の間のリンクも提供します。
言語: 今すぐ一緒に
言語は、上で説明した多くの脳領域の調整された機能を活用します。言語には、さまざまな音声、さまざまな文字、およびそれらの組み合わせを区別できる、非常に洗練された知覚能力が必要です(なじみのない外国語を聞いたり見たりするだけで、自分の母国語の知覚能力がどれほど素晴らしいかを体験できますが、これは当然のことだと思っています)。同様に、言語の音を出し、文字や単語を書くためには、高度なモーター出力能力が必要です。その中間で、言語は、多くの場合高速で流れてくる文法的および意味論的な情報を追跡するために、最も要求の厳しい形式の認知処理を必要とします。これには、すべての意味情報を統合するための強力な分散後皮質意味表現に加えて、高度な実行機能と作業記憶能力が必要です。
言語 章での探求は、私たちの脳が単語の意味をどのように表現するかを考えることから始まります。私たちは、自己組織化ネットワークが 単語の共起における統計的規則性をエンコードする方法を学習し、単語 [@LandauerDumais97] の類似構造を捉えるのに非常に効果的な 意味論的 表現を生み出す方法を探ります。このネットワークは、このテキストの初版の初期草案に基づいてトレーニングされるため、関連するセマンティクスをよく理解している必要があります。
次に、正書法 (ライティング) から 音韻論 (スピーチ) への経路のモデルに目を向け、このスペルと音のマッピングにおける規則性と例外に関する問題を調査します。これは、[@SeidenbergMcClelland89; @PinkerPrince88; @PlautMcClellandSeidenbergEtAl96] でかなりの議論のテーマとなっています。知覚の章のオブジェクト認識モデルには、通常のマッピングと例外のマッピングの両方をサポートする機能の重要な組み合わせがあることを示し、このモデルは非単語プローブ入力を人間と同じように発音し、英語のマッピング構造に関する同様の基礎知識を抽出したことを示します。
次に、これら最初の 2 つのモデルのコンポーネントを、個々の単語の正書法、音韻論、および意味論的な表現を相互接続する小規模な読解モデルにまとめます。これらのコンポーネントは 分散辞書 を形成します。すべての単語情報が 1 か所に保存されるわけではありません。代わりに、関連する知覚、運動、および意味論的な情報の処理に特化した脳領域全体に分散されます。興味深いことに、このモデルの特定の経路に損傷を与えることで、さまざまな形態の 後天性失読症 (例: 脳卒中や他の形態の脳損傷による) をシミュレートすることができ、侵襲的な実験が不可能な人間の言語機能の神経相関関係を確立する重要な方法を提供します。
最後に、文ゲシュタルト 表現の概念を使用して、文の意味を処理するコンテキストでの構文と意味論の間の相互作用に取り組みます。この表現では、粗くコーディングされた分散表現を使用して文全体の意味をエンコードし、構文と意味論の両方の手がかり [@StJohnMcClelland90] を統合します。これは、言語理論でよく使用される記号的で高度に構造化されたアプローチとは対照的に、構文に対する明らかに神経的なアプローチです。