compcogneuro/book: メモリ
このページは外部資料の日本語訳です。原文の見出し順と本文順を保ち、コード・URL・出典表記はできるだけ原形のまま残しています。
出典とライセンス
原典: https://github.com/compcogneuro/book/blob/main/chapter-07.md
ライセンス: CC BY 4.0。このページは日本語翻訳であり、変更点は翻訳とサイト内整形です。
bibfile: ccnlab.bib —
メモリ
記憶について考えるとき、おそらくエピソード記憶、つまり特定のエピソードや出来事の記憶を思い浮かべるでしょう。もしかしたら、子供の頃の特別な出来事(誕生日、家族旅行など)や、トラウマになった出来事(スーパーマーケットで道に迷ったり、ハイキングや家族での外出中に置き去りにされたことはありますか?)を覚えているかもしれません。おそらく、昨日の夕食に何を食べたか、誰と食べたか覚えているでしょうか?記憶のこの側面は私たちにとって最も顕著ですが、それはさまざまな種類の記憶の 1 つにすぎません。
機械的、計算的な用語での記憶の大まかな区分の 1 つは、重みベース と 活性化ベース の形式の記憶です。体重に基づく記憶はシナプス可塑性の結果であり、一般に比較的長く持続します(少なくとも数十分、場合によっては数十年、最大で一生)。活性化ベースの記憶は進行中の神経活動によってサポートされているため、はるかに一時的でつかの間ですが、より柔軟でもあります。重みに基づく記憶は脳内のすべての変更可能なシナプスに存在するため、無数の方法で現れる可能性があります。この章では、心理学者によって研究された記憶の最も顕著なタイプのいくつかに焦点を当てます。まずエピソード記憶から始めて、親密度に基づく認識記憶、次に重みに基づくプライミング、および活性化に基づくプライミングを見ていきます。 実行機能 の章では、作業記憶を含む、より堅牢な形式の活性化ベースの記憶について見ていきます。
おそらく、ほとんどの人は 海馬 と、エピソード記憶におけるその重要な役割について聞いたことがあるでしょう。たとえば、映画 メメント は、海馬が機能しないことがどのようなものかを見事に描写しています。私たちは計算モデルを通じて、なぜ海馬がエピソード記憶に優れているのかを明らかにします。海馬は神経活動の非常にまばらなパターン(一度に活動するニューロンが比較的少ない)を持っているため、比較的似た記憶であっても、非常に異なる重複しない神経表現を持つことができます。これらの独特の神経パターンは、記憶の主な敵である「干渉」を劇的に軽減します。実際、大脳新皮質の高度に分散され、重複する表現は、本書の前半で概説した理由から有用ではあるものの、あまりに急速に学習しようとすると、それ自体で壊滅的な干渉を引き起こします。しかし、エピソード記憶に必要なのは、この迅速な「ワンショット」学習です。その代わりに、脳は 2 つの特殊な **相補的な学習システム ** を活用しているようです。新しいエピソード記憶を迅速に符号化する海馬と、意味論的知識の豊富な網をゆっくりと取得する新皮質です。これらは、後で説明するように、重複する分散学習と遅い学習速度から大きな恩恵を受けます。
脳を過剰に単純化してモジュール化しようとする常に存在するように見える衝動に対抗するには、記憶は高度に分散された現象であり、脳全体の何十億ものシナプスがあらゆる特定の経験によって微調整されることを理解することが重要です。いくつかの研究では、海馬に重大な損傷を負った人々において、比較的特定の情報に関する新しい記憶の学習が保存されていることが示されているが、これらの記憶がどのように手がかりとなるかを考慮することが重要である。これはメモリ全般について覚えておくべき重要な側面です。特定のメモリを実際に取得できるかどうかは、システムのプローブ方法に大きく依存します。おそらく誰もが、昔のたまり場を訪れた結果、大量の記憶が甦るという経験をしたことがあるのではないでしょうか。利用可能な無数の手がかりによって、そうでなければ表面に浮上するほど強力ではなかった記憶が(一見自発的に)呼び戻されるのです。海馬の恩恵を受けずにエンコードされた記憶は弱く、より曖昧ですが、確かに存在します。
脳内の記憶は高度に分散されていることに加えて、高度にインタラクティブでもあります。最初に脳のある部分でエンコードされた情報は、それらの記憶が再活性化され、これらの他の脳領域がそれを学習するさらなる機会を得た場合、脳の他の部分に「広がった」ように見えることがあります。典型的な例は、海馬で最初にコード化されたエピソード記憶が、それらの記憶を繰り返し呼び出すことによって周囲の新皮質領域で強化される可能性があるということです。これは、日中に体験した記憶のパターンが再び活性化されることがわかっている睡眠中にも起こる可能性があります。さらに、前頭前皮質システムと感情状態の影響は、記憶の符号化と検索に大きな影響を与える可能性があります。したがって、コンピューターの静的な「ハードドライブ」の比喩とは程遠く、脳内の記憶は非常に動的であり、脳のすべての領域に存在する複雑さと相互作用を反映し、常に進化するプロセスです。
エピソード記憶
私たちはエピソード記憶から始めます。それは、それが私たちの意識生活、そして実際に私たちのアイデンティティの非常に重要な部分を占めているからです。たとえば、映画 トータル リコール は、フィリップ K. ディックの小説 私たちはあなたのためにそれを覚えておくことができます卸売 (Wikipedia リンク) に大まかに基づいており、エピソード記憶と自己意識との関係を探求しています。海馬が機能している人は皆、この驚くべき「テープレコーダー」を持っており、起きている間に起こるすべてのことを常にエンコードしています。20分前や数時間前に起こったことを思い出すのに特別な努力をする必要はありません。それはただ自動的にそこにあります。ほとんどの人は、日常生活の移り変わりの大部分を結局忘れてしまい、特に顕著な、または意味のある出来事だけを保持します。
しかし、一見「普通」に見えるごく一部の人々は、異常に多数の経験を鮮明に詳細に思い出すことができます (これは 気分亢進症 (Wikipedia へのリンク) と呼ばれます)。興味深いことに、これらの人々を区別するのは海馬自体ではありません。代わりに、彼らは強迫的なリハーサルとエピソード記憶の想起を特徴とし、大脳基底核の領域が明らかに拡大しています(これは強迫性障害(OCD)に関連しています)。 実行機能 の章で説明するように、大脳基底核は運動制御と強化学習だけでなく、活動記憶の更新と維持の強化にも関与しています。これは、正常な人間の脳では、海馬が私たちの生活の毎日を暗号化して記憶する生の能力を持っていることを示唆していますが、ほとんどの人は、それらの記憶をすべて確実に取り出せるレベルまでリハーサルすることを気にしません。実際、気分過敏症の人が抱える主な不満は、ほとんどの人が手放してしまう人生の不快なことをすべて忘れることができないことです。
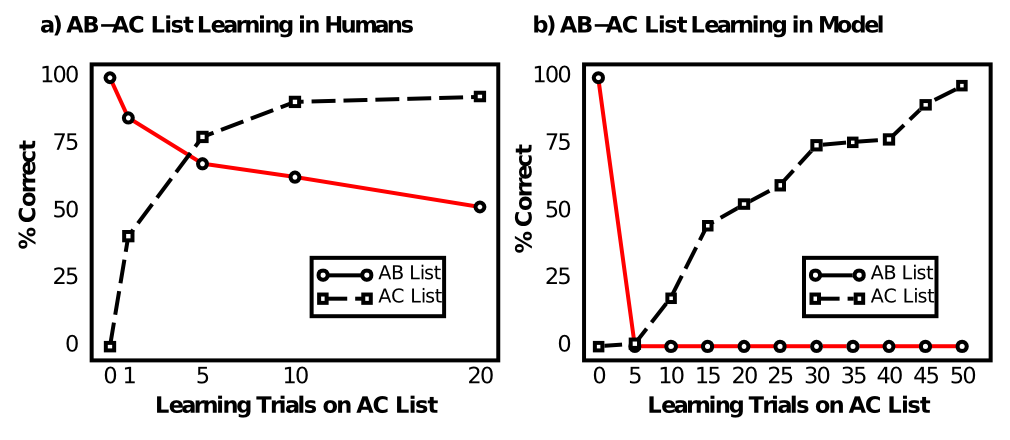
それでは、海馬が非常に優れたエピソード記憶システムである理由は一体何でしょうか?私たちの調査は失敗から始まります。具体的には、この教科書で私たちが検討してきた種類の「一般的な」皮質ニューラルネットワークモデルが、何らかの有用なエピソード記憶能力を発揮できないことです。この障害は、AB-AC ペア対応リスト学習タスク ([@fig:fig-ab-ac-list]) で訓練された汎用バックプロパゲーション ネットワークを使用して、[@McCloskeyCohen89] によって初めて文書化されました。このタスクには、AB リストと呼ばれる、任意の単語のペアの初期リストを学習することが含まれます。例:
*機関車 - ふきん
- ウィンドウ - 理由 ・自転車・木
- …
人々は、各 A 項目に関連する B を思い出す能力がテストされ、AB リストでのトレーニングは完全に思い出すことができた時点で終了します。次に、以前の A 項目の新しい仲間を含む AC リストの学習を開始します。
- 機関車 - 雲
- ウィンドウ - 本
- 自転車 - ソファ
- …
この AC リストの学習を 1、5、10、20 回繰り返した後、人々は元の AB 項目について追加のトレーニングを行わずに、元の AB 項目を思い出す能力がテストされます。左側のプロットは、AC 項目を学習した結果、2 つのリスト間にかなりの重複があるため、AB リストにかなりの量の干渉があることを示していますが、AC 項目を 20 回繰り返した後でも、人々は依然として AB リストの約 50% を思い出すことができます。対照的に、右側のプロットは、ネットワーク モデルが 壊滅的な干渉を示し、AB リストのパフォーマンスが即座に 0% になったことを示しています。彼らは、明らかに人間の方がはるかに優れたエピソード記憶能力を持っているため、これによって人間の認知に関するすべてのニューラルネットワークモデルが無効になると結論付けました。
しかし、この種のニューラル ネットワークの全面的な放棄は正当化されないことがわかります (実際、脳は巨大なニューラル ネットワークであるため、あらゆる現象について何らかのニューラル ネットワークの記述が必要であり、私たちはこの種の課題を、関連するメカニズムを特定するための有益な機会として捉えています)。実際、次の調査では、干渉のレベルを軽減する特定のネットワーク パラメータがあることがわかります。必要な最も重要な操作は、抑制レベルを上げてアクティブなニューロンを減らすことです。これにより、AB リスト項目と AC リスト項目の内部表現間の重複が減少し、システムが以前の AB メモリを上書きすることなく AC を学習できるようになります。次に、海馬システムがこのトリックを (他のいくつかのトリックとともに) 極端に活用し、非常に優れたエピソード記憶システムになっていることがわかります。
壊滅的な干渉の調査
CCNシムズ から abac シミュレーションを実行します。
海馬とパターンの分離/パターンの完成
海馬は、(パターン分離による) 重複と干渉を最小限に抑える高度にまばらな表現(つまり、アクティブなニューロンが比較的少ない)を使用してエピソード記憶を迅速に記録するように特に最適化されています。この考えは非常に大量のデータと一致しており、確立された事実に近いものです(認知神経科学では珍しいことです)。このデータには、海馬の選択的病変に起因するエピソード記憶障害(特にパターン分離)に関する基本的な所見、他の脳領域と比べて説明を求めるほど独特な海馬の解剖学的特徴、およびさまざまな海馬領域からの神経記録データの膨大なレパートリーが含まれています。まず海馬の解剖学的構造の概要を説明し、続いて神経記録データと、比較的まばらな神経活動レベルがどのようにしてパターン分離を引き起こし、干渉を最小限に抑えるかを理解します。
海馬の解剖学
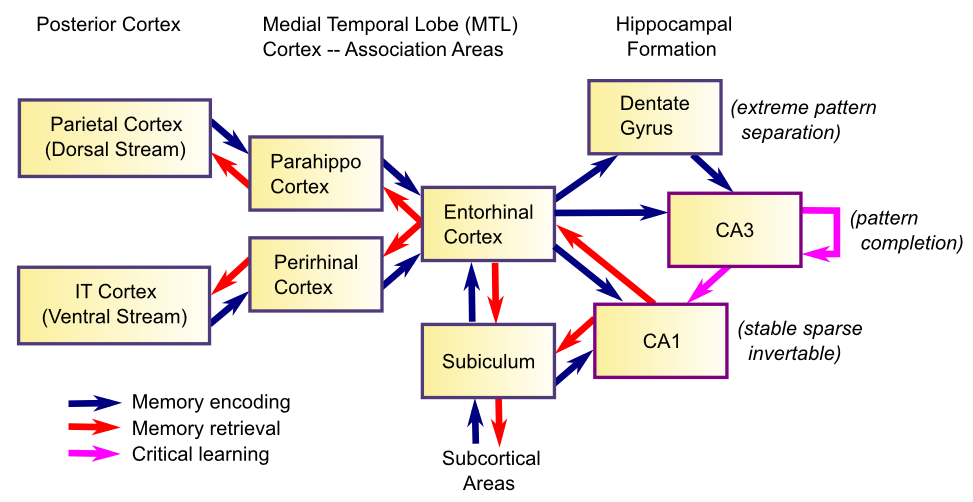
海馬自体の解剖学と海馬に栄養を与える領域は [@fig:fig-hippo_mem_formation-7] に示されています。海馬は、相互に接続された皮質領域の階層の最上部にある 2 つの「頂点」の 1 つを表します (最下位は感覚入力領域、たとえば一次視覚野です)。もう 1 つのそのような頂点は、実行機能 の章で検討される前頭前野です。したがって、これはエピソード記憶システムにとって重要な機能を備えています。つまり、その時点で脳内で起こっている注目すべきすべての非常に高レベルの要約にアクセスできるのです。この情報は、知覚と注意 の章で検討した背側経路と腹側経路の二重経路に沿って整理され、海馬傍 (PHC) (背側) 領域と 鼻周囲 (PRC) (腹側) 領域に収束し、嗅内皮質 (EC) に送られ、その後海馬本体に送られます。海馬の主な領域には、歯状回 (DG) と「アンモンの角」 (ラテン語でコルヌ アンモニス (CA))、CA3、CA1 の領域が含まれます (CA2 に何が起こったのでしょうか? 基本的に CA3 と同じであることが判明したため、そのラベルを使用するだけです)。これらの奇妙な名前はすべて、人間の脳にあるタツノオトシゴの形を指す「海馬」という用語自体を含め、これらの領域の形状に関係しています(海馬はギリシャ語でタツノオトシゴを意味します)。
この構造に関する基本的なエピソード記憶エンコーディングのストーリーは次のようになります。脳内のあらゆるものの高レベルの要約が EC で活性化され、穿孔経路 を介して DG および CA3 領域を駆動します。この最終結果は、海馬の主要な「エングラム」を表す、非常にまばらで明確な神経発火パターンである CA3 です。 EC は CA1 のアクティビティも駆動します。CA1 には、この同じ EC パターンをすべて単独で再アクティブ化できるという重要な機能があります (つまり、CA1 と EC の間の 反転マッピング または 自動エンコーダー 関係)。これらの活動パターンは、相互接続されたすべてのシナプスのシナプス可塑性 (学習) を促進します。最も重要なのは、CA3 ニューロン間のシナプス接続 (CA3 回帰経路)、および CA3 と CA1 の間の接続 (シェイファー側副経路) です。これらの可塑的変化は、CA3 エングラム内のさまざまなニューロンを効果的に「接着」し、それらを CA1 の可逆パターンと関連付けます。そのため、その後の CA3 エングラムの検索により、CA1、次に EC が活性化され、皮質に戻ることができます。したがって、海馬の主な機能は、エピソードのすべての異なる要素を結合し、この 結合記憶 を取り出して、想起中に皮質に復元できるようにすることです。これが、記憶が「フラッディングバック」する仕組みです。CA3 から CA1、EC、皮質へとフラフラと戻ってきて、記憶がエンコードされた時点の元の脳パターンに近い何かを再活性化します。
冒頭で述べたように、この方法でメモリを単純化してモジュール化しようとする試みはすべて不正確であり、実際、メモリのエンコードはエピソード時にアクティブなすべてのニューロンに分散されます。たとえば、パーフォラント経路での学習は、EC 入力から CA3 エングラムを再活性化するために重要です (特に、EC 入力が部分的な記憶想起手がかりのみを表す場合)。さらに、海馬に出入りする皮質経路全体を学習することで、検索プロセスが「スムーズに」行われます。実際、記憶パターンが頻繁に再活性化される場合、これらの皮質結合は、海馬の恩恵をまったく受けずに、完全な記憶の再活性化を促進するのに十分強力である可能性があります。この 統合 プロセスについては後で詳しく説明します。最後に、前頭前野を使用したトップダウン戦略を使用して記憶を制御して検索することによって、検索プロセスを強化できます。ここでは制御された取得のこの側面については考慮しませんが、実行機能 の章で検討するいくつかの機能と同様の、アクティベーションと重みベースのメモリの組み合わせに依存します。
海馬ニューロンの特性: 疎性、パターン分離
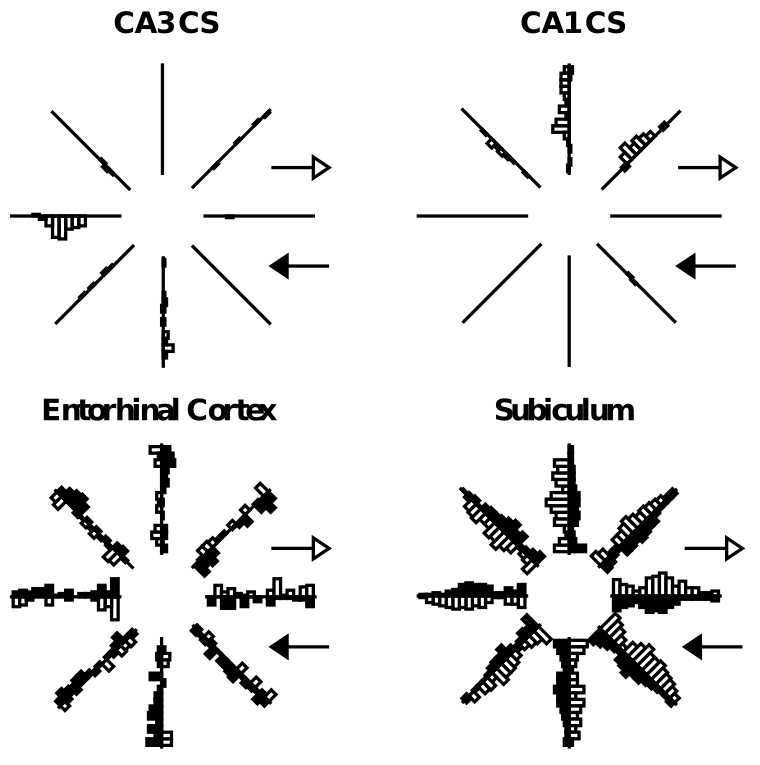
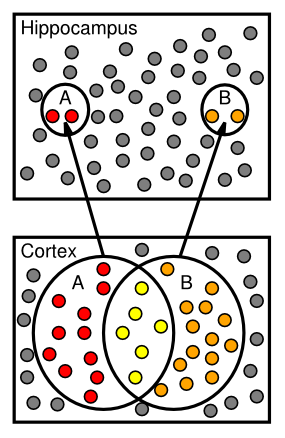
海馬 (CA3、CA1) と皮質の間の決定的な違いの代表的な写真が [@fig:fig-hcmp-rfs] に示されています。ここでは、CA3 および CA1 ニューロンの発火頻度が皮質 (嗅内皮質および海馬台) のニューロンよりもはるかに低いことが明らかです。これが、海馬表現における まばらさ の意味です。どのエピソードでも、比較的少数のニューロンだけが発火し、逆に、各ニューロンは非常に特殊な状況下でのみ発火します。ラットでは、これらの状況は空間的な位置、つまり 場所細胞 として識別される傾向がありますが、これは霊長類の海馬には一般に当てはまりません。このまばらさは、これらの領域での高レベルの GABA 阻害によって生じると考えられており、多くのニューロンが閾値以下に保たれ、この阻害を克服するには活動的なニューロンが比較的高レベルの興奮性入力を受け取る必要があります。このまばらさの直接的な利点は、基本的な確率 ([@fig:fig-patsep-clr]) から見て、異なるエピソードのエングラムの重複が少なくなることです。たとえば、特定のエピソードでニューロンがアクティブになる確率が 1% (DG の典型) である場合、任意の 2 つのランダムなエピソードの確率は、その値の 2 乗、つまり 0.01% (非常に小さな数) になります。比較すると、確率がより高い場合、たとえば 25% (皮質に典型的)、2 つのエピソードの重複の可能性は 6.25% になります。 David Marr は、影響力のある論文 [@Marr71] の中で、スパース表現のこの「パターン分離」特性を最初に指摘したようです。

活動レベルとパターン分離の間の関係は、DG ニューロンと CA3 ニューロンの発火特性を比較することによって、海馬自体の内部でも観察できます。DG ニューロンは、ややまばらな CA3 (活動レベル約 2 ~ 5%) と比較しても、活動レベルが最もまばらです。 [@LeutgebLeutgebMoserEtAl07] による研究からの [@fig:fig-leutgeb-et-al-07-dg-patsep] は、正方形から円へ、そして再び円へ戻る環境の系統的なモーフィングの関数として、DG が CA3 よりも多くのパターン分離を示すことを示しています。 DG ニューロンは、この環境変化に応じてより多様な神経発火を示し、これらの異なる環境を CA3 よりも大幅に分離していることを示唆しています。他にも、皮質と比較したさまざまな海馬領域、特に他の領域と比較した DG におけるパターン分離の説得力のある実証が数多くあります (たとえば、このテーマに関する Kesner の広範な研究を参照)。
効果的なパターン分離に寄与するもう 1 つの要因は、穿孔経路を介した EC から DG および CA3 への広範囲かつ拡散した接続です。これにより、EC のさまざまな機能を DG と CA3 でランダムに組み合わせることができ、入力の組み合わせまたは 結合 に敏感になることができます。まばらな活性化に関連する高い抑制閾値のため、これは、これらの領域の特定のニューロンがこれらの拡散入力ソースの複数から顕著な興奮を受け取る必要があることを意味します。言い換えれば、これらのニューロンは結合表現を持っています。
関係するニューロンのパターンの重複が比較的少ないため、パターンの分離は、事前の学習への干渉を最小限に抑えながら、海馬が新しいエピソードを迅速にエンコードできるようにするために重要です。
パターンの完成: キューのリコール
新しい記憶をエンコードするにはパターン分離が重要ですが、これらの記憶が後で呼び戻されない限り、このエンコードは役に立ちません。この呼び出しプロセスは パターン完了 としても知られており、部分的な検索キューが記憶に関連付けられた完全なオリジナル パターンの完了をトリガーします。たとえば、「子供の頃にサマーキャンプに行きましたか?」という質問をしたとします。場合によっては、ここからサマーキャンプの思い出をパターン化することも、そうでないこともできます。人間の記憶の驚くべき点は、それが内容参照可能な記憶であることです。十分に特定された情報のサブセットが検索の手がかりとして機能し、以前にエンコードされたエピソード記憶の回復が可能になります。対照的に、コンピュータのメモリはメモリ アドレスまたは変数ポインタによってアクセスされ、メモリに格納されている実際の内容とは何の関係もありません。 Google などの最新の Web 検索エンジンは、コンテンツのアドレス指定可能性の重要性を実証しており、人間の記憶システムとよく似た機能を持ち、検索語を検索の手がかりとして利用して、関連情報を含む関連する「記憶」(Web ページ) を見つけます。 Web を検索することでご存知かと思いますが、クエリを具体的にすればするほど、関連する情報を取得できる可能性が高くなります。同じ原理が人間の記憶にも当てはまります。
海馬では、CA3 ニューロン間の反復的な接続によってパターンの完成が促進され、エンコード中に CA3 ニューロンが結合され、CA3 ニューロンのサブセットが残りの部分の想起を引き起こすことができます。さらに、穿孔経路におけるエンコード中のシナプスの変化により、元の DG および CA3 ニューロンが部分的な検索キューによって再活性化される可能性が高くなります。
興味深いことに、パターンの分離とパターンの完成の間には直接的な緊張関係またはトレードオフがあり、海馬の解剖学的構造の詳細なパラメーターがこのトレードオフ [@OReillyMcClelland94] を最適化していると見なすことができます。パターン分離により、システムが検索キューを新しい刺激のように扱い、古いエングラム パターンを完成させるのではなく、新しい別個のエングラム パターンを CA3 にエンコードする可能性が高くなります。同様に、システムがパターン補完に優れている場合、真に斬新なエピソードを実現するために、新しいパターンを分離したものをエンコードする代わりに、古い記憶を再アクティブ化します。私たちのモデルの解剖学的パラメーターは、これらの異なる完了力と分離力の間の良好なバランスを見つけるのに役立ちますが、海馬は、現在の要求がそれぞれ想起を必要としているかエンコードを必要としているかに応じて、完了または分離のいずれかを強調するために、前頭前皮質実行制御領域などの他の脳領域からの戦略的影響の恩恵を受けている可能性もあります。この問題については、執行機能の章でさらに詳しく説明します。
探検
AB-AC タスクを使用して、海馬がどのように記憶をエンコードして呼び出すかを調べるには、CCNシムズ で hip シミュレーションを実行します。
補完的な学習システム
前に述べたように、マクロスキーとコーエンが壊滅的な干渉の現象を初めて発見したとき、彼らはニューラル ネットワークには致命的な欠陥があり、人間の認知の実行可能なモデルと見なすべきではないと結論付けました。これは、隠れ層がないため XOR などのより難しいマッピングを学習できないネットワークのコンテキストで、[@MinskyPapert69] で起こったのと同じことです (詳細については、学習 の章を参照してください)。どちらの場合も、これらの問題に対する解決策はすでに用意されていますが、人々は心のニューラル ネットワーク モデルを軽視する言い訳を好んで利用するようです。おそらく、脳内で起こっているすべてが実際に単なるニューロンに集約される可能性があると考えるのは、あまりにも還元主義的であるか、そうでなければ恐ろしいです…しかし、この問題はニューラルネットワークに特有のものではないかもしれません—研究者は、データのあるパターンと一致しない場合、たとえばベイジアンモデルを含む心のさまざまな理論を無視することがよくあります。重要なのは、困難なデータを考慮して、特定の理論に根本的な欠陥があるかどうかを特定することです。悪魔は細部に宿っており、多くの場合、「赤ちゃんを風呂のお湯と一緒に捨てる」ことなく、既存の理論を調整したり改良したりする方法があります。
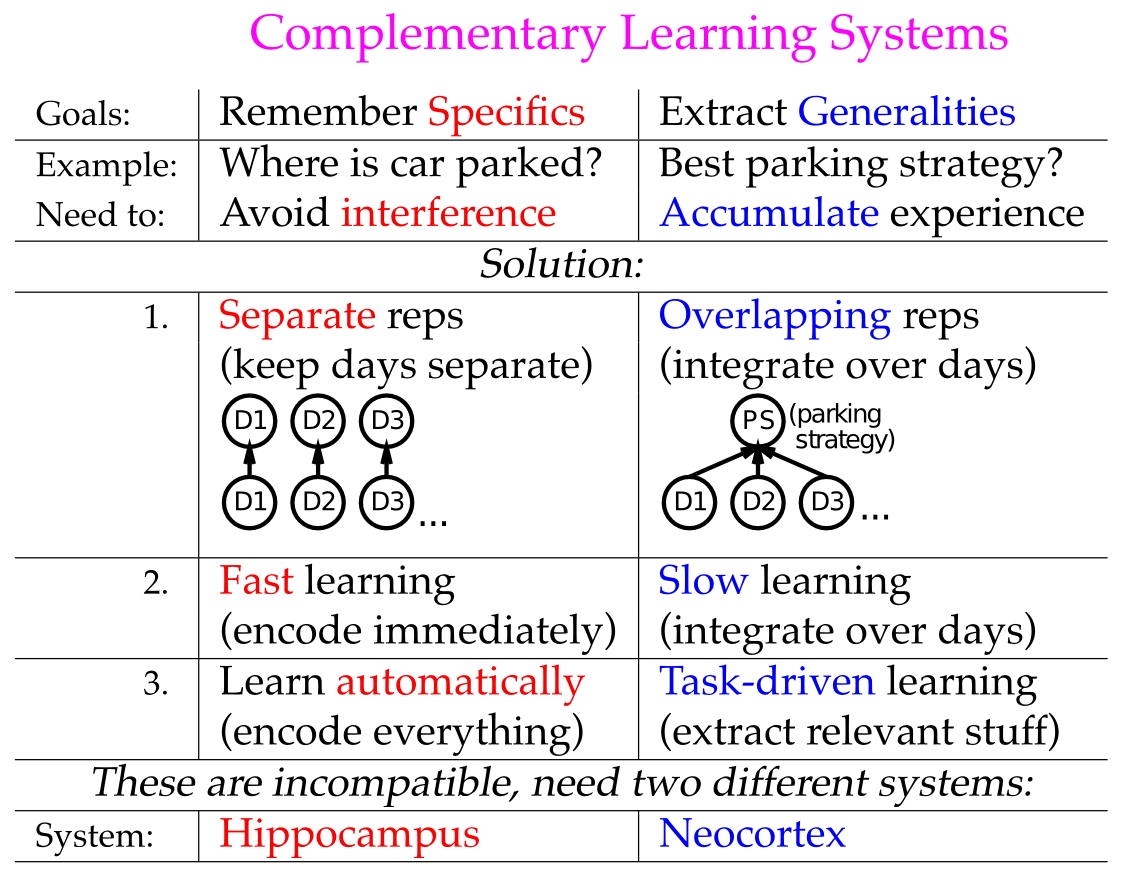
このような考えはさておき、壊滅的な干渉問題には (少なくとも) 2 つの解決策が考えられます。 1 つは、必然的に、形成される表現間の重複を何らかの形で減らすことによって、エピソード記憶タスクにおける汎用ニューラル ネットワーク モデルのパフォーマンスを何らかの方法で改善することです。もう 1 つは、特殊なエピソード記憶システム、つまり海馬を導入することです。海馬は、新皮質学習のモデルとして一般的なニューラル ネットワークの機能を保持しながら、パターン分離による低干渉の迅速な学習のために特に最適化されたパラメーターを備えています。 相補学習システム (CLS) フレームワーク [@McClellandMcNaughtonOReilly95; @NormanOReilly03] として知られるこの後者の視点の利点は、一般的な神経モデルのエピソード記憶を改善するために行うことが、実際には新皮質の優れたモデルとなる能力を妨げることです。
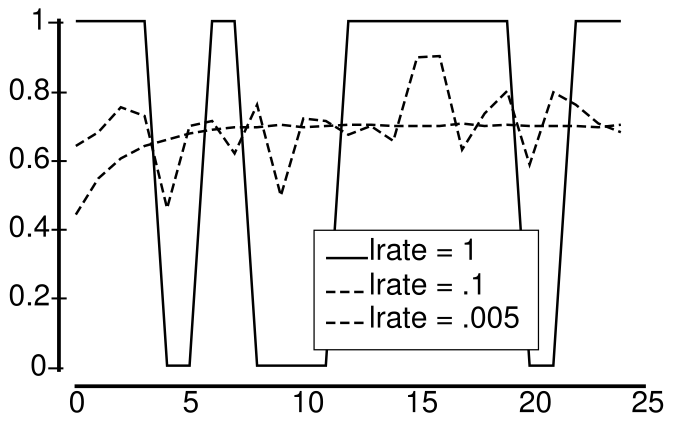
具体的には、物体認識 (知覚と注意 の章で説明したように) や意味論的推論 (言語 の章で説明したように) などの新皮質学習は、高度に重複する分散表現と低速のインターリーブ学習から恩恵を受けます。これらの重なり合う分散表現により、神経活動のパターンが項目 (物体、単語など) 間の複雑な高次元の類似構造をコード化できるようになります。これは、世界の「常識的」理解を得るために重要です。 [@fig:fig-hippo-comp-learn-sys] は、統計的学習または意味学習 (新皮質に関連) とエピソード記憶 (海馬に関連) の間の基本的なトレードオフを要約しています。 [@fig:fig-integ-lrate] は、特定の結果の全体的な確率を計算するための離散エクスペリエンス全体の統合における学習率の影響を示しています。この統合を適切に実行できるのは、より速い学習率で発生するようにごく最近の過去を反映するだけではなく、遅い学習率だけです。
この基本的なトレードオフと一致して、(前述したように) 優れたエピソード記憶能力を持つ人々は、エピソード間で知識を一般化することに相応の困難に悩まされることがよくあります。さらに極端な、さまざまなトピックに関するあらゆる詳細な情報を記憶できる自閉症の記憶力の高い人は、一般に、現実の世界で生きていくための常識的な推論と一般的な能力がさらに深刻に欠如しています。これらの場合、新皮質も海馬によく似た機能を持ち、活動パターンがまばらになり、その結果、全体的に詳細を記憶する能力が向上しますが、それに応じて経験全体を一般化し、常識的な推論 [@McClelland00] を生み出す能力が低下すると推測されます。
健忘症: 順行性と逆行性
無傷の海馬がどのように機能するかを見たので、健忘症を引き起こす何が問題なのか疑問に思うかもしれません。ハリウッド版の健忘症では、頭を殴られ、その後、知っていることすべてを完全に忘れてしまいます(たとえば、配偶者が他人になるなど)。その後、もちろん、別の良い攻撃でそれらの記憶が復元されますが、その後、多くのおかしな悪事が続きます。実際には、健忘症にはさまざまな原因があり、記憶の研究者は通常、海馬健忘症として知られる、海馬および関連構造への直接的な損傷によって引き起こされる種類に焦点を当てています。この最も有名な症例は、科学界でH.M. として知られている人物です。 (ヘンリー・モレゾン)、彼は難治性てんかんを予防するために海馬を切除しました。その後、新しいエピソード情報を学習できなくなる (前向性健忘) だけでなく、以前に学んだ知識をある程度忘れてしまう (逆行性健忘) ようになりました。しかし、彼は話し方、さまざまな言葉や物の意味、自転車の乗り方を覚えており、あらゆる種類の新しい運動能力を学ぶことができました。これは、海馬が特定の種類の新しい知識のみを学習するために重要であることを明確に示しました。
H.M.とさらに注意深く研究してください。彼は新しい意味論的な情報を学習することもできるが、その学習は比較的ゆっくりと行われ、神経学的に無傷の人々に比べて、学習した知識にアクセスする方法がより脆弱であることを示した。これは、海馬がエピソード学習にとって重要であるが、意味論的学習には重要ではないことをさらに明確にします。ただし、ほとんどの人にとって、意味情報は最初は海馬を介して学習され、その後時間をかけて新皮質によってよりゆっくりと取得されます。このプロセスが起こっていることを示す 1 つの兆候は、H.M.彼は、古い記憶(つまり、時間的に段階的に変化する逆行性勾配、リボット勾配としても知られる)よりも、手術前の最新の記憶を失っています。したがって、古い記憶はどういうわけか海馬の外で「統合」されており、最初に海馬でエンコードされた新皮質学習情報のこの段階的なプロセスが実際に起こっていることを示唆しています。このプロセスについては次のセクションで説明します。
特定の薬剤は、選択的な前向性健忘症を引き起こす可能性があります。たとえば、ベンゾジアゼピン (広く研究されている薬剤 * ミダゾラム * を含む) は脳全体の GABA 抑制ニューロンを活性化しますが、ベンゾジアゼペン (GABA-A) 受容体は海馬で高密度に発現しており、抑制レベルが高いため、これに対して非常に敏感です。適切な用量であれば、この阻害は海馬内でシナプス可塑性が生じて新しい記憶が形成されるのを防ぐのに十分ですが、以前に学習した記憶が再活性化される可能性はあります。これにより、逆行性のない、より純粋な順行性健忘症が生じます。実験的には、ミダゾラムは海馬依存の高速記憶エンコードを阻害しますが、強化学習 [@HirshmanPassannanteArndt01; @FrankOReillyCurran06] などの他の形式の統合学習を妨げます。
健忘症の別の原因はコルサコフ症候群に由来し、通常は長期のアルコール依存症によるビタミン B1 の欠乏によって引き起こされます。これは明らかに視床と乳頭体の一部に影響を及ぼし、次に内側中隔からのGABA神経支配を含むさまざまな神経調節経路を介して海馬に影響を及ぼし、海馬の学習と想起のダイナミクスに影響を与える可能性があります。
海馬から新皮質への記憶の統合
なぜ私たちは夢を見るのでしょうか?私たちが眠っている間に私たちの脳の中で何か有益なことが起こっているのでしょうか、それともそれは単なるランダムなノイズとごちゃ混ぜの無意味な連想なのでしょうか?実際に寝ながら外国語を学べるのでしょうか?睡眠と夢の謎に対する私たちの絶え間ない魅力は、睡眠中に記憶が何らかの形で海馬から新皮質に移動するという考えをめぐる興奮を説明できるかもしれません。 記憶の固定として知られるこのプロセスは、当初、H.M. の場合のように、人が後天性健忘症に苦しんでいる場合、より最近の記憶が失われる可能性が高いという観察によって動機づけられました。上で説明しました。さらに最近では、覚醒中と睡眠中の海馬の神経記録により、ラットが迷路を走っているときに起こる活動パターンが、ラットが眠っているときにも再活性化されるようであることが明らかになった。ただし、測定された再活性化のレベルは、実際の行動中に活性化されたパターンと比較して比較的弱いため、そこからどの程度強い学習信号が生成されるかは明らかではありません。さらに、適切に制御された動物実験における時間的に段階的な逆行勾配の存在についてはかなりの論争があり、そもそも圧密現象の存在についていくつかの疑問が生じています。それにもかかわらず、全体的に見て、このプロセスは、完全に遍在していなくても、少なくともある程度、少なくともいくつかの状況で発生すると結論付けるのが安全であるように思われます。人間の場合、ノンレム睡眠中の徐波振動は記憶の固定と関連していると考えられています。実際、ある最近の研究では、睡眠中に徐波振動を外部から誘導すると、睡眠[@MarshallHelgadottirMolleEtAl06]の直前にコード化された項目について、その後の海馬依存性の記憶が実際に強化されることが示されました。
この統合プロセスに関する相補的学習システムの観点からの予測の 1 つは、これら 2 つのシステムにおける学習と表現の性質が大きく異なるため、新皮質でエンコードされた情報は海馬で最初にエンコードされた情報とは異なる性質を持つということです。したがって、エピソード記憶が新皮質でエンコードできる限り、もともと海馬でエンコードされていたより明確で鮮明なパターン分離表現と比較して、エピソード記憶はより「意味化」され、他の既存の記憶と統合されて一般化されるでしょう。利用可能な証拠は、例えば、海馬健忘症からの無傷の記憶の性質を神経学的に無傷の対照と比較することによって、この考えを支持しているようである。
海馬における空間の役割
海馬に関する多くの研究はラットで行われており、空間ナビゲーションはラットにとって最も重要な行動機能の 1 つです。したがって、ラットの海馬が強力な 場所細胞 発火 ([@fig:fig-hcmp-rfs] に示すように) を示し、個々の DG、CA3、および CA1 ニューロンが空間内の特定の位置に応答することは、おそらくそれほど驚くべきことではありません。特定のニューロンは、異なる環境では異なる場所細胞の位置を持ち、これらの場所細胞にはいかなる種類の地形やその他の体系的な組織もないようです。これは、これらの領域への穿孔経路の投影のランダムで拡散した性質、およびパターン分離の効果と一致します。
さらに最近では、グリッド細胞の形で、嗅内皮質の空間コーディングが発見されました。これらのグリッドセルは空間上に正六角形の格子またはグリッドを形成し、さまざまな形の振動に依存しているように見えます。これらのグリッド セルは、海馬本体内の場所セルに統合される生の空間情報を提供する可能性があります。さらに、頭方向細胞が海馬に突き出たさまざまな領域で発見されており、これらの細胞は、最近の動きの蓄積に基づいて、ラットがどこを向いているかについて優れた推測航法信号を提供します。
これらすべての細胞型の組み合わせは、ラットの空間ナビゲーションに強固な基盤を提供しており、これらの異なる信号がどのように連携してナビゲーション行動をサポートできるかを示すさまざまな計算モデルが開発されています。このドメインの探索モデルは、将来のエディションで利用可能になる予定です。
シータ波
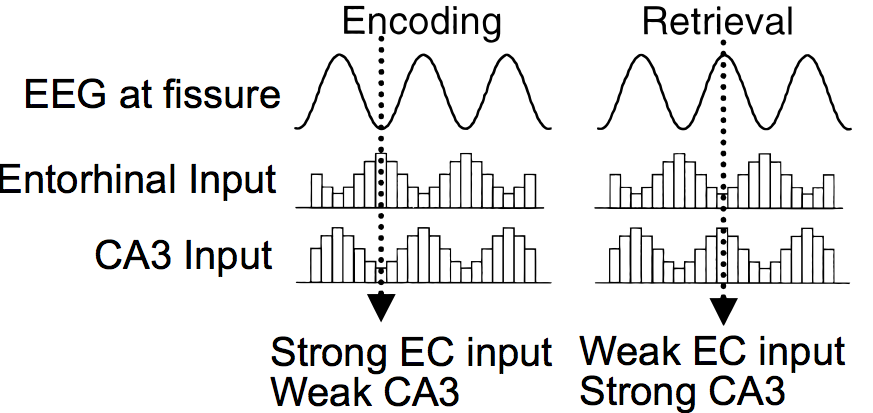
海馬の重要な特性は、ラットのいわゆる *シータ * 周波数帯域における神経発火率の全体的な振動であり、その範囲は 1 秒あたり約 8 ~ 12 回です。 [@fig:fig-hasselmo-bodelon-wyble-theta] に示されているように、海馬のさまざまな領域はこのシータ振動に関して互いに位相がずれており、これらの位相差により海馬がより効果的に学習できる可能性が生じます。 Hasselmoらは、このシータ位相の関係により、システムは新しい情報の符号化と既存の情報[@HasselmoBodelonWyble02]の再現を交互に行うことができると主張した。これは魅力的なアイデアです。なぜなら、前に説明したように、海馬パラメータを変更して、他のさまざまな種類の要求に基づいてエンコードまたは検索を最適化することで利点が得られるからです。
Emergent ソフトウェアは、この基本的なシータ エンコーディングと検索のアイデアの拡張をサポートするようになりました。これにより、Leabra エラー駆動学習が、[@KetzMorkondaOReilly13] を処理する 1 つの標準トライアル内で、海馬における 2 つの異なる学習経路を形成できるようになります。各経路には、有効なマイナス相とプラス相の活性化状態があります (実際には、同じプラス相を共有します)。標準的なマイナスからプラスへの位相差で訓練された主経路には、対応する CA1 活動パターンの CA3 駆動の想起が含まれ、EC などを再活性化することができます。 2 番目の経路は、マイナス相内での安定という特別な初期相を使用して訓練され、CA1 <-> 経路です。 EC 可逆自動エンコーダ。EC が正しくリコールされた場合、CA1 が実際に EC を再アクティブ化できることを保証します。以前に調査した標準的な海馬モデルでは、この自動エンコーダー経路は、EC および CA1 ユニットの単一のサブグループ (「スロット」と呼びます) 内のすべての可能なサブパターンで事前にトレーニングされています。この新しいモデルは、この自動エンコーダーがシータ位相サイクルを介してどのように学習できるかを示唆しています。 この海馬の シータ期 バージョンの詳細については、付録 海馬シータ期 を参照してください。
シータ振動は、EC 層のグリッド セルの活性化にも重要な役割を果たしていると考えられており、おそらく、場所フィールドの活動の発火が シータ段階の行列 を示し、展開するシータ波内の異なる点で異なる場所フィールドが発火するため、時間シーケンス情報をエンコードする役割も果たしている可能性があります。これらのトピックについては、今後の改訂版でさらに詳しく説明します。
海馬台の機能
海馬台は海馬機能の理論では無視されることが多いですが、それでもさまざまな重要な役割を果たしている可能性があります。解剖学的には、他の海馬領域と比較して嗅内皮質 (EC) と同様の位置に位置していますが、新皮質領域と相互接続される代わりに、皮質下領域とより直接的に相互接続されています ([@fig:fig-hippo_mem_formation-7])。したがって、EC と類推すると、EC は海馬への皮質下情報の入出力経路と考えることができます。海馬台が実行する可能性のある非常に重要な機能の 1 つは、特定の状況の相対的な新規性を計算し、これを中脳のドーパミン システム、さらには大脳基底核に伝達して、行動を適切に調節することです。目新しさは複雑な感情的な結果をもたらす可能性があり、不安誘発性(不安を引き起こす)とさらなる探索を促す動機付けの両方をもたらし、一般的に全体的な覚醒レベルを高めます。海馬は、関連する空間情報やその他の文脈情報の完全な「結合」を考慮して、状況がどの程度新規であるかを独自に判断することができます。たとえば海馬台は、シータ振動の想起フェーズ中に CA1 と EC の状態を比較することで新規性を計算できる可能性がありますが、これは現時点ではまったくの推測です。この斬新な信号を組み込むことは、将来の計算モデルの重要な目標です。
親しみやすさと認識記憶
ここで、海馬の特定の記憶への貢献から一歩下がって、海馬システムが人間の記憶容量のより大きな空間にどのように適合するかについて、より広い視点から検討します。研究者たちがここで焦点を当てた最も重要な疑問の一つは、新皮質が単一試験のエピソード記憶に何らかの寄与をすることができるかどうかである。与えられた刺激への 1 回の曝露は、明白な行動に影響を与えるほど大きな痕跡を皮質のどこかに残すのでしょうか?前述したように、脳全体のシナプスがあらゆる学習経験によって影響を受ける可能性が高いと私たちは確信していますが、単一の経験から行動的に重要な変化を生み出すには、新皮質の学習が単に遅すぎたり、表現が重複しすぎたりするのでしょうか?
大量のデータは、確かに新皮質はエピソード記憶の痕跡をサポートできるが、それらは海馬によってサポートされるものと比較して非常に異なる特性を持っていることを示唆しています。具体的には、鼻周囲皮質は、特定の刺激が最近経験されたかどうかを非常に大まかに示す、有用な「慣れ」信号を生成できるようです。この親近感信号は、海馬によって提供される回想記憶信号、つまり、そのアイテムが最後に経験されたときの前のエピソードの詳細を完全に明示的に思い出す信号と対比することができます。親密度シグナルはむしろ、アイテムの親密度の強さに応じて強度が変化する単一の段階的な値に似ています。この信号の神経基盤に関する仮説の 1 つは、鼻周囲皮質における表現の「鮮明さ」です。一般的な皮質モデルでの学習を 1 回試行すると、神経活動の全体的なパターンに測定可能な痕跡が残り、強く活動するニューロンとより弱く活動するニューロンの間のコントラストが強調されます (Norman & O’Reilly、2003)。これは、学習 章で観察した基本的な自己組織化学習ダイナミクスの結果であり、最も強く活性化されたニューロンがシナプス接続を強化するため、他のニューロンとよりよく競争できるようになります。
興味深いことに、人々はこの親近感信号に主観的に意識的にアクセスし、それを利用して、そのアイテムがどれだけ親近感を持っているかについての明白で意識的な評価を行うことができます。シャープネス信号をこのように明示的に読み出すための神経機構はまだ解明されていません。ここでの主な課題は、なぜ鼻周囲皮質の信号が意識的にアクセスできるのに、他の新皮質領域の同様の信号は意識にアクセスできないように見えるのかを特定することです(次のセクションで説明します)。
海馬の想起と鼻周囲の精通記憶システムのこの組み合わせは、認識記憶の「二重プロセス」モデルと呼ばれ、長年の論争を経て、現在ではこの分野で広く受け入れられています。この二重プロセスモデルと一致するデータの一部には、海馬に大きな損傷を負った人々の保存された親密度信号や、これら 2 つの記憶信号をさまざまな方法で区別することができたさまざまな神経画像および行動研究が含まれます。
プライミング: 重みとアクティベーションベース
海馬および周囲の皮質領域(例えば、鼻周囲皮質の慣れ信号)からさらに離れたところに移動すると、知覚およびその他の連合皮質領域は、1回または少数の曝露に基づいて有用な記憶に貢献できるでしょうか?ここでの答えも肯定的ですが、親近感の信号とは異なり、これらの記憶の痕跡は意識のレーダーの下にほぼ完全に残ります。科学者はさまざまな行動尺度の観点から記憶の効果を測定できますが、私たちはこれらの記憶があることを主観的に認識していません。この形式の記憶の一般的な用語は プライミング です。主な行動の現れは、反応時間の高速化、または特定の行動反応を起こす確率の増加であるためです。あたかもこれらの記憶の痕跡によって「ポンプが呼び水されている」かのようです。実際、私たちは、新皮質のゆっくりとした漸進的な学習効果が、まさにこのポンプの呼び水レベルの調整を根底にある神経表現に対して行っていると考えています。多くの経験にわたる持続的な変化のみが、これらのより安定した神経表現をより劇的な方法で真に再形成することができます。そして、年齢を重ねるにつれて、おそらく学習速度が遅くなり、最も基本的な新皮質の表現を根本的に再形成することがさらに困難になっているようです。
遅い学習変化による微妙な影響に加えて、プライミングは残留活性化、つまり以前に処理された情報から持続する神経発火によっても生じる可能性があります。したがって、重みベースのプライミングとアクティベーションベースのプライミングを区別できます。予想されるとおり、活性化ベースのプライミングは非常に短期間であり、神経発火が消えるとすぐに消えます。対照的に、重量ベースのプライミングは非常に持続性が高く、1 回の曝露でプライミングが 1 年以上続くケースもあります。この種の行動結果は、シナプス可塑性の安定性に強い制約を課します。さまざまな計算モデルではシナプス重量減衰の形態が導入されていますが、これはプライミングの極度の耐久性、そしてより一般的には私たちの長期記憶と矛盾しているようです。
プライミング効果を明らかにするために使用される行動パラダイムの 1 つは、ステム補完 と呼ばれます。ここでは、単語の最初の文字が提示され、参加者は最初に思いついた単語で語幹を完成させるように求められます。たとえば、次のようなステムが表示される場合があります。
*勝ち___ *__させてください
「窓」や「冬」、「手紙」や「レタス」などの言葉で答えます。プライミング効果は、最初に人々にこれらの語幹の考えられる単語の 1 つを、多くの場合かなり偽装された偶発的な方法で曝露し、次にそれがその後その語幹を完成させる可能性にどの程度影響するかを比較することによって明らかにされます。人々がどの単語にさらされるかをランダム化することで、人々がどのようなベースラインの好みを持っているかに関連して、以前にさらされた言葉の影響を分離することができます。これらのプライミング効果は海馬での学習によるものではないことがわかっています。なぜなら、海馬に病変がある人でもプライミング効果はそのまま残っているからです。
探検
CCNシムズ の priming シミュレーションでは、非常に汎用的な皮質学習モデルを使用して、単純なステム完了のようなタスクで重みベースと活性化ベースのプライミングの両方を探索できます。
## 付録
- 海馬シータ期: 海馬のシータ期学習バージョン。
海馬シータ期
この付録では、hip 探索で使用されるシータ期海馬実装 [@KetzMorkondaOReilly13] に関する詳細情報を提供します。 ソース コードと現在の実装の詳細については、github の リーブラ/ヒップ を参照してください。
ネットワーク内のアクティベーション ダイナミクスの 3 つのフェーズを次に示します。
-
マイナスフェーズの前半: EC 入力層が CA1 を駆動し、CA1 が EC 出力層を駆動します。CA1 は CA3 の影響を受けません (シータサイクルでは CA3 は抑制されますが、実際には CA1 に影響を与える有効な重みスケールを 0 に設定しているだけです)。これは、EC <-> CA1 自動エンコーダー経路をトレーニングするためのマイナスフェーズです。
-
マイナスフェーズの後半: CA3 は CA1 に影響を与えてリコールを駆動しますが、EC 入力は CA1 を駆動しません。安定化の終わりのこの状態は通常のマイナスフェーズであり、プラスフェーズと比較して学習を促進し、CA3 -> CA1 想起経路を訓練します。
-
プラスフェーズ: EC 出力層ユニットは EC 入力層のアクティビティによって直接アクティブ化され、EC 出力は EC 入力パターンを再現することを学習します。CA1 もマイナスフェーズの前半と同じ状態にあり、CA3 ではなく EC 入力によって駆動されます。したがって、CA3 -> CA1 リコール接続のターゲット CA1 アクティビティ パターンは、EC 出力層で EC 入力を適切にリコールする状態であり、同様に、この同じパターンが EC <-> CA1 オート エンコーダのターゲットになります。