compcogneuro/book: 実行機能
このページは外部資料の日本語訳です。原文の見出し順と本文順を保ち、コード・URL・出典表記はできるだけ原形のまま残しています。
出典とライセンス
原典: https://github.com/compcogneuro/book/blob/main/chapter-09.md
ライセンス: CC BY 4.0。このページは日本語翻訳であり、変更点は翻訳とサイト内整形です。
bibfile: ccnlab.bib —
実行機能
私たちは現在、認知神経科学の階層の最上位である「幹部」レベルに到達しています。ビジネスにおいて、経営者は、会社のさまざまな部門から入ってくる高度な情報に基づいて、「収益」を強く考慮して、重要な意思決定や計画を立てます。人間の場合、主に 前頭前皮質 (PFC) 内で行われると考えられている実行レベルの処理は、同様に後部皮質連合野から高レベルの情報を受け取り、最終的に行動を導く「最終的な」力を伝える動機付けおよび感情の領域とも直接相互接続されています。私たちの多くは、自分の行動が合理的な思考と計画に基づいているという印象(妄想?)を持って歩き回っていますが、むしろ、基本的な生物学的動機と感情信号が私たちの行動を形作る上で重要な役割を果たしている可能性が非常に高いです。少なくとも、これは PFC および関連する脳領域の基礎となる生物学が示唆していることです。それでもなお、PFC は、特定の目標を追求する際に気を散らすものやその他の影響を無視する能力など、より抽象的な推論と計画能力をサポートするために重要であることも明らかです。この章では、PFC におけるこの一見矛盾した能力の共存の謎を解き明かしていきます。
高次レベルの認知制御における PFC の重要性の証拠は、PFC の損傷に関連する 環境依存症候群 から得られます。典型的な例では、PFC 損傷のある患者が研究者の家を訪れ、ベッドを見るとすぐに服を脱ぎ(カツラも外して!)、ベッドに入り、寝る準備をしました。環境の手がかりは、あまりよく知らない人の家で何をすべきかについてのこれまでの文脈を圧倒しました。言い換えれば、PFC がなければ、行動はより抽象的で熟考された計画や目標ではなく、直接的な感覚環境のアフォーダンスによって動かされ、はるかに反射的で思考力のないものになります。この症候群を経験するのに実際の PFC 損傷は必要ありません。確かに、あなたも、それ以外の場合は計画していなかった直接の感覚環境に触発されて、ぼんやりと何かをしているという経験をしたことがあります (たとえば、たまたま歯ブラシが見えたため、就寝前に 2 回歯を磨くなど)。私たちは皆、注意力の低下を経験します。ぼんやりとした教授という古典的な固定観念は、教授に PFC が欠如していることで説明されるのではなく、むしろ PFC が明らかに別のことに取り組んでおり、そのため脳の残りの部分が環境に依存して自分自身で対処できるようにしているからです。
PFC の認知への貢献についての洞察を得るもう 1 つの優れた情報源は、私たち一人一人が毎晩、夢の形で得ることができます。 PFC は、夢を見ている睡眠中に最も不活性化される脳領域の 1 つであることが判明しました。その結果、私たちの夢はしばしば連続性を欠き、最も接した糸だけがそれらを繋いでいる状態で、ある切り離された場面から別の場面に飛び移っているように見えます。たとえば、ある瞬間には高校時代の緊張した社会状況を追体験しているかもしれません。そして次の瞬間には、飛行機の出発時刻を調べようとしていて、どうしようもなく遅れるのではないかという恐怖を感じているかもしれません。
では、PFC が頭脳の幹部として機能できる独自の理由は何でしょうか?答えの一部は、上でほのめかしたように、その接続性です。脳の情報処理階層全体の最上位に位置し、脳内で起こっているすべての重要なことについて、高度に処理された「ステータス レポート」を受け取ります。この意味では、記憶の章で見た海馬に似ており、実際、これらの領域は連携して機能しているようです。しかし、PFC はまた、私たちの行動を制御するのに特に適した位置にあります。PFC は前頭運動野の直前にあり (運動 の章を参照)、明白な (および隠れた) 運動行動を駆動する広範な接続性を備えています。さらに、PFC の内側領域と腹側領域は、扁桃体などの皮質下領域の感情処理領域と直接相互接続されているため、PFC が動機付けおよび感情信号によって駆動され、相互に増幅または無効化されることが可能です。
PFC は、適切な場所に存在するだけでなく、入力される感覚信号などの注意散漫に直面しても情報を保持できるようにするいくつかの特別な生物学的特性も備えています。したがって、PFC が正常であれば、他人のベッドに横になるという考えに抵抗し、訪問の目的に集中し続けることができます。この能力は、目標やその他の関連情報を維持するために必要な期間にわたってニューロンの集団をアクティブに発火し続ける能力に依存するため、この能力を 堅牢なアクティブ メンテナンスと呼びます。この能力は 作業記憶 とも呼ばれますが、この後者の用語は文献の中でさまざまな方法で使用されているため、この文脈では PFC における情報の堅牢でアクティブな維持と同義であると慎重に定義します。アクティブ メンテナンスがゲーティング システムとどのように連携するかについては後で説明します。これにより、一度に複数の項目を念頭に置き、一部の情報を選択的に更新および操作しながら他の情報を維持し続けることができ、統合システムがより洗練された形式の作業記憶をサポートできるようになります。
1970 年代のサルの PFC のニューロンの記録では、サルが遅延を超えて活発に発火する (別名 遅延期間活動) ことを示しました。最も広く使用されている課題の 1 つは、眼球運動遅延応答課題です。この課題では、ビデオ ディスプレイの特定の位置で刺激がフラッシュされますが、サルは中央の固視十字に目の焦点を合わせ続けるように訓練されており、その十字が消えた時点でジュース報酬を受け取るために、以前にフラッシュされた位置に目を移動する必要があります。前頭眼野(PFCの領域)のニューロンは、刺激の位置に同調した強力な遅延周期発火を示し、この活動は、サルが遅延後に正しく目を動かした直後に終了します。人間の PFC におけるこの堅牢な能動的な維持については、他にも多くの実証があります。
この章で検討する計算モデルは、接続性と堅牢なアクティブ メンテナンスのこれら 2 つの要素がどのように組み合わされて、PFC に起因する幅広い実行機能能力をサポートできるかを示します。目標は、サポートすると考えられている認知能力の膨大なリストと比較して、実行機能の統一モデルを提供することです。
最も重要な実行機能能力の 1 つは、行動や思考を戦略的に 迅速に変更する能力 (認知的柔軟性と呼ばれることが多い) です。たとえば、パズルやその他の難しい問題を解決しようとするとき、多くの場合、適切な解決策を見つける前に、さまざまなアイデアを試す必要があります。 PFC がなければ、行動は反復的で常同的なもの (何度も何度も壁に頭をぶつける) となり、この特徴的な柔軟性が欠けています。 PFC でアクティブに維持されている内容を迅速に更新する機能により、PFC システムは行動や思考を迅速に変更できます。システムの動作を変更するために比較的ゆっくりとしたシナプス重みの変更を必要とする代わりに、PFC でアクティブな神経発火のパターンを更新することで、動作を即座に変更できます。つまり、PFC システムは活性化状態を動的に更新することで行動の適応に貢献し、関連する活動パターンの トップダウン バイアスを通じて後部皮質表現や運動動作を形成します。対照的に、後皮質または大脳基底核における行動の適応は、体重の変化にゆっくりと適応することに大きく依存します。この違いの証拠は、広く研究されている成人の ウィスコンシン カード ソーティング タスク (WCST) や子供の * 次元変化カード ソーティング タスク (DCCS)* などのタスク切り替えパラダイムから得られます。
この章の計算モデルは、大脳基底核 (BG) と 中脳ドーパミン領域 (特に 腹側被蓋野、VTA) が PFC 活動状態の迅速かつ戦略的な更新においてどのように重要な役割を果たしているかを示します。具体的には、堅牢なアクティブ メンテナンスには、既存の情報の維持と新しい情報をエンコードする更新を切り替える追加の制御信号が必要であることがわかります。 BG は、おそらく VTA からのドーパミン作動性信号と連携して、PFC における情報維持の 動的ゲート の役割を果たします。この動的なゲート機能は、モーター の章で見たように、モーター動作のゲート制御において BG が果たす役割と同じです。さらに、BG の学習プロセスも、強化学習の原理に基づいたモーターの章の学習プロセスと同じです。具体的には、(VTAの隣にあるSNcからの)ドーパミンがBG学習を形成し、それによってゲーティングメカニズムが、維持することが重要なもの(タスクに関連しており、したがって本質的報酬を予測できるもの)と無視できるもの(タスクの良好なパフォーマンスを予測できないため)を決定するという困難な問題に対処できるようにします。これらのメカニズムは、PFC-BG 認知システムが、運動行動と学習を制御するための既存の強力なメカニズムを活用することによって進化したという一般的な概念を具体化しています。この観点から見ると、ドーパミン作動性学習シグナルは PFC/BG 機能の基本機構において中心的かつ密接な役割を果たしているため、認知はモチベーションから切り離すことはできません。認知および運動行動の選択と学習におけるBGとドーパミンの同様の機能は、サルの研究からの証拠や、ヒトにおけるBGおよび/またはドーパミンに影響を与える疾患の影響、薬理学的操作、機能的画像処理、遺伝学など、過去10年または20年にわたるさまざまなデータによって強力に裏付けられてきました。
強力なアクティブメンテナンスをサポートする PFC/BG とドーパミンの生物学
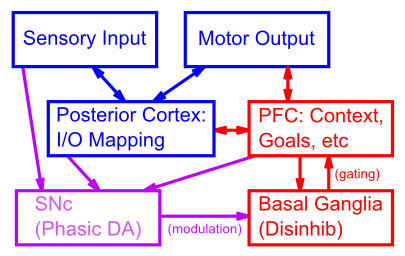
実行機能にとって特に重要な領域の全体的な接続性は、後皮質 (側頭葉、頭頂葉、後頭葉) と運動前頭皮質 (つまり、前頭前皮質の後側の前頭皮質) に関連する感覚および運動処理に関連して、[@fig:fig-pbwm-architecture-bio] に示されています。 PFC は、後皮質の高次連合皮質領域と相互接続されており、感覚世界に関する高度に処理され抽象化された情報が符号化されています。また、上位レベルの運動制御領域(運動前野、補足運動野)と相互接続し、下位レベルの運動制御信号を調整して、調整された一連の運動出力を実行します。この接続パターンにより、PFC は後部皮質と運動皮質で行われる処理から受信し、影響を与えることができる位置にあります。
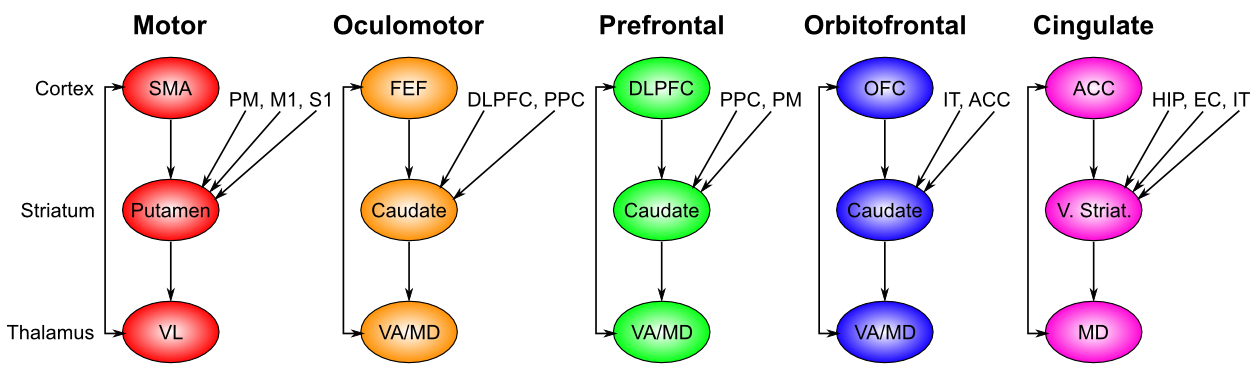
大脳基底核 (BG) は、主に線条体 (尾状核、被殻、側坐核)、淡蒼球、視床下核から構成され、視床の特定の核 ([@fig:fig-bg_loops_ads86-9]) を介して PFC と密に相互接続されています。 運動制御と強化学習 の章で詳しく説明されているように、BG は、PFC と視床間の興奮性ループを脱抑制することにより、前頭葉皮質に動的で適応的なゲート制御の影響を与えます。 PFC のコンテキストでは、このゲートの影響は、モーター制御のコンテキストでモーター動作の開始を制御するのと同じメカニズムを使用して、PFC 内でアクティブに維持される情報の更新を制御します。また、PFCから視床下核へのトップダウンの投射は、進行中の行動選択を停止または切り替えるべき条件を検出し、BG回路の残りの部分が計画された運動行動をゲートするのを防ぐことにより、行動に対する一種の抑制制御をサポートします。
実行制御システムの最後の主要な構成要素は、黒質緻密部 (SNc) と他のいくつかの関連脳領域で構成され、これらが一緒になって BG の相性ドーパミン神経調節を駆動し、そのゲート動作の強化学習をもたらします。このシステムは、モーター制御と強化学習 の章で説明されているように PVLV モデルを使用して計算的に要約されており、PFC 内の情報のアクティブな維持と相互作用して、BG 内のゲート信号を強化することができ、その後の良好なパフォーマンスと報酬につながります。段階的 DA 強化学習のこのタイムトラベル特性は、時間の経過とともに情報を維持するシステムをトレーニングするために不可欠です。
以下のサブセクションでは、これらの各システムの生物学的特性と実行機能との関連性を要約します。 「A-not-B」課題のモデルで、積極的な維持能力の発達が乳児の不可解な行動をどのように説明できるかを探っていきます。ストループ タスクのモデルが、堅牢なアクティブ メンテナンスの観点から、実行機能 (および機能不全) の非常に幅広い側面をどのように捉えているかを見ていきます。どの情報を積極的に維持すべきかをシステムがどのように決定できるかを調べるために、前頭前皮質大脳基底核作業記憶モデル (PBWM) に目を向けます。PBWM は、大脳基底核と中脳のドーパミン領域を統合して、複雑な実行機能タスクを実行します。
PFC での堅牢なアクティブ メンテナンス
![持続的な遅延期間のアクティビティの例 (パネル B)。遅延サッカード課題中に前頭眼野 (FEF) で記録された個々の細胞の活動率のヒストグラム (背景のドット) と曲線。ターゲット刺激は、試験の開始時に短時間だけオンになります(パネル A、ターゲット、メモリ)。この細胞は、他の細胞が試験の終了時に正しいサッカードを生成できるように、遅延中にその活動を維持した(パネルD、E)。 [@SommerWurtz00]、図 2 から転載。](https://raw.githubusercontent.com/compcogneuro/book/main/figures/fig_SommerWurtz00_fig2.png)
[@FusterAlexander71; @KubotaNiki71] によって最初に発見された、遅延を超えて持続的な能動発火を示す PFC ニューロンの能力は、序文で説明した遅延サッケーディング課題との関連で、[@fig:fig-SommerWurtz00-fig2] のパネル B (「遅延信号を持つニューロン」) に示されています。 PFC ニューロンの他のサブセットも、視覚入力に一時的に反応したり (パネル C)、運動を開始したり (パネル D) するなど、他の発火パターンを示します。 PFC における神経応答パターンのこの区別には、後で説明する PBWM モデルで捉える重要な機能的意味があります。
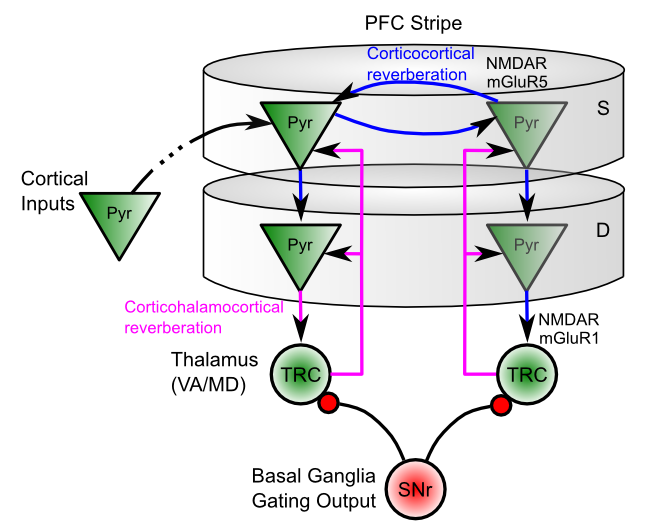
PFC ニューロンが時間の経過とともに持続的に活動的な発火を示すことを可能にする 2 つの主要な生物学的メカニズムがあります。
-
反復性興奮性接続: PFC ニューロンの集団は強力な興奮性相互接続 ([@fig:fig-pfc-trc-reverb-loops]) を持っており、その結果、神経発火がこれらの相互接続されたニューロン間で前後に反響し、結果として持続的な活性発火が生じます。このような接続には 2 つのタイプがあります: 1) 同じ PFC ストライプ内の錐体細胞間の 皮質皮質ループ、および; 2) PFC の VI 層錐体細胞と、その特定の細胞グループに投影する視床中継細胞との間の 皮質視床皮質ループ。
-
固有の興奮性維持電流: 反復性興奮性ループの両方によって形成されるシナプスには、NMDA 受容体と代謝指向性グルタミン酸 (mGluR) 受容体があり、高周波活動によって一度開放されると、残響活動を継続させるために興奮性が増大するより長い時間枠が提供されます。 学習 の章を思い出してください。NMDA チャネルでは、チャネルをブロックする Mg+ (マグネシウム) イオンを除去するためにニューロンが十分に脱分極される必要があります。 NMDA チャネルのこの活動依存の性質により、NMDA チャネルは、「スイッチ」または動的にゲートされた形式のアクティブな維持を提供するのに理想的に適しています。すでに十分に活性化されているニューロンのみが、これらの NMDA チャネルによって提供される興奮の増加から恩恵を受けることができます。これは、大脳基底核システムが能動的な維持を制御するための「フック」を提供します。視床ニューロンが BG ゲート作用によって抑制解除されると、その後の活動のバーストにより、PFC ニューロンのサブセットが NMDA Mg+ ブロック閾値を超え、それによって時間の経過とともに強力に発火し続けることが可能になります。
PFC アクティブ メモリ強度の開発と A-not-B タスク
発達プロセスは、多くの場合、認知の失敗を特に明確にし、関連するプロセスに光を当てることによって、さまざまな認知現象に対する重要な洞察を提供します。この好例は、先駆的な発達研究者のジャン・ピアジェ [@Piaget54] によって開発された A-not-B タスクです。幼児は、ある場所 (A というラベル) に隠されたおもちゃを繰り返し見せられ、その後そのおもちゃが別の場所 (B) に隠されると、引き続き A に手を伸ばします。その行動は印象的です - 幼児はおもちゃが隠されているのを見たばかりで、実験者の動きを細心の注意を払って追跡しています(通常、斬新で興味深いおもちゃが使用されます)。それにもかかわらず、彼らはこのことをすべてすぐに忘れて、以前に確立された「習慣的な」行動に戻ってしまうようです。この課題の成功は、背外側前頭前皮質領域 [@Diamond90] の発達と完全性にかかっています。
ここで検討する計算モデル [@Munakata98] は、非常に微妙で複雑なものも含め、一連の動作現象を、この章の後半で検討するストループ モデルと多くの共通点を共有する比較的単純なモデルでどのように捉えることができるかを示しています。このモデルにおける発達は、PFC ニューロン間の残響興奮性結合の強さとして単純に操作可能であり、これがこの単純化されたモデルにおける能動的な維持のための唯一のメカニズムです。 「古い」ネットワークは、再帰接続が強いため、情報 (おもちゃがどこに隠されたかなど) を長期間保持できますが、再帰接続が弱い「若い」ネットワークでは、情報ははるかに一時的です。
アクティブ メンテナンスの開発の検討
これがどのように展開されるかを確認するには、CCNシムズ で a-not-b モデルを開きます。
PFC 領域にわたる機能の特化
堅牢なアクティブ メンテナンスのメカニズムは PFC 全体に存在しますが、異なる PFC 領域は全体的な実行機能への異なる貢献に関連付けられています。この章の後半では、これらのさまざまな機能的寄与が、すべての PFC 領域が重要な機能として堅牢なアクティブメンテナンスを共有するという統一された枠組みの中で、これらの PFC 領域と脳の他の部分との接続性の違いという観点から説明できるという考えを検討します。
![前頭前皮質の領域のブロードマン数。それぞれの領域は実行機能の異なる混合に関連付けられています。 [@Fuster01] より転載。](https://raw.githubusercontent.com/compcogneuro/book/main/figures/fig_Fuster01_2.png)
解剖学的には、前頭葉は中央溝の前の皮質領域を構成します。中心溝のすぐ前、したがって前頭皮質の最も後方にあるのは一次運動野 (M1) です。これは外側の表面で最も顕著に見られますが、背側の表面を越えて内側まで広がっています。 M1 のほぼ前方にある連続組織は、計画運動野、運動前野 (PM) 皮質 (外側)、および補足運動野 (SMA、SMA 前; 内側) を構成します。次に、その前に PFC エリアがあり、[@fig:fig-Fuster01-2] のブロードマン番号でラベルが付けられています。
最も広いレベルでは、PFC 領域は内側対外側、背側対腹側という主軸に沿って分割できます。一般に、外側PFC野は、より後皮質の感覚野および運動野と相互接続されており、これらの領域の処理を制御する役割を果たしていると考えられています。対照的に、内側 PFC 領域は、感情機能や動機付け機能に関連する皮質下の脳領域とより強く相互接続されています。機能的には、外側領域は「コールド」認知制御に重要である一方、内側領域は「ホット」な感情および動機の処理 [@fig:fig-cortical_fun-9] に重要であると特徴付けることができます。ただし、この区別は思ったほど明確ではありません。側面領域でさえ、維持された認知情報がタスクの成功(報酬の一種)を予測する程度に基づいて、動機付け変数やBG/ドーパミンゲート信号によって調整されるからです。
![後部皮質の *何を* 対 *どのように* の区別を前頭前皮質に持ち込んで、PFC の腹側領域と背側領域の独特の役割を理解することができます。 [@OReilly10] から転載。](https://raw.githubusercontent.com/compcogneuro/book/main/figures/fig_cortical_fun_org_tins_acc_ofc.png)
背側と腹側の区別の機能的重要性については、文献においてかなり議論の余地があるが、解剖学的には、背側 PFC 野が後皮質の背側経路とより相互接続しているのに対し、腹側 PFC は腹側後皮質経路と相互接続していることは明らかである。 知覚 の章で見たように、後皮質の背側経路は、運動制御を駆動するために知覚信号を抽出する、行動に対する知覚 (どのように 処理) に特化しているのに対し、腹側経路は識別のための知覚 (何を 処理) に特化しています。この後皮質の機能的特殊化は、PFC [[@fig:fig-cortical_fun-9]] の関連する背側野と腹側野に引き継がれる可能性があります。そのため、背側外側 PFC (DLPFC) 野は、運動計画の実行制御と運動制御を駆動する頭頂皮質経路の制御に特に重要であり、腹側外側 PFC (VLPFC) 野は、体の存在を識別する側頭葉経路の制御に特に重要です。また、これらのエンティティに関する豊富な意味上の関連付けも形成します。
内側では、背側内側PFCは前帯状皮質(ACC)としても知られており、運動制御変数の感情的な側面(たとえば、行動にどれだけの労力がかかるか、成功の確率はどれくらいか、反応を選択する際にどの程度の葛藤と不確実性があるかなど)をコード化することが示されており、これは「ホット・ハウ」機能の専門化と一致しています。背内側 PFC 領域は BG 内の視床下核にも投影し、難しい応答選択要求 [@Frank06; @AronBehrensSmithEtAl07; @CavanaghWieckiCohenEtAl11] の下での衝動的な選択を防ぐために運動応答を遅らせる役割を果たします。眼窩前頭皮質 (OFC) を含む PFC の腹内側領域 (VMPFC) は、さまざまな感覚刺激の感情値をコード化していることが示されており、これらの領域が「熱い」領域であるという考えと一致しています。 What/How 機能特化のアイデアの詳細については、[@OReilly10] を参照してください。
持続的な PFC 発火によるトップダウンの認知制御: ストループ モデル
ここで、実行機能のテストに使用されるおそらく最も研究されているタスクである ストループ タスクに目を向けます。これは、基本的な現象 [@Stroop35] を最初に説明したジョン リドリー ストループにちなんで名付けられました。 [@CohenDunbarMcClelland90] によって最初に開発されたこのタスクの計算モデルは、(ラベルを適切に変更して) 非常に広範囲のさまざまな現象に適用されています。したがって、この一見単純なタスクとモデルは、実行機能の最も重要な特徴を捉えています。

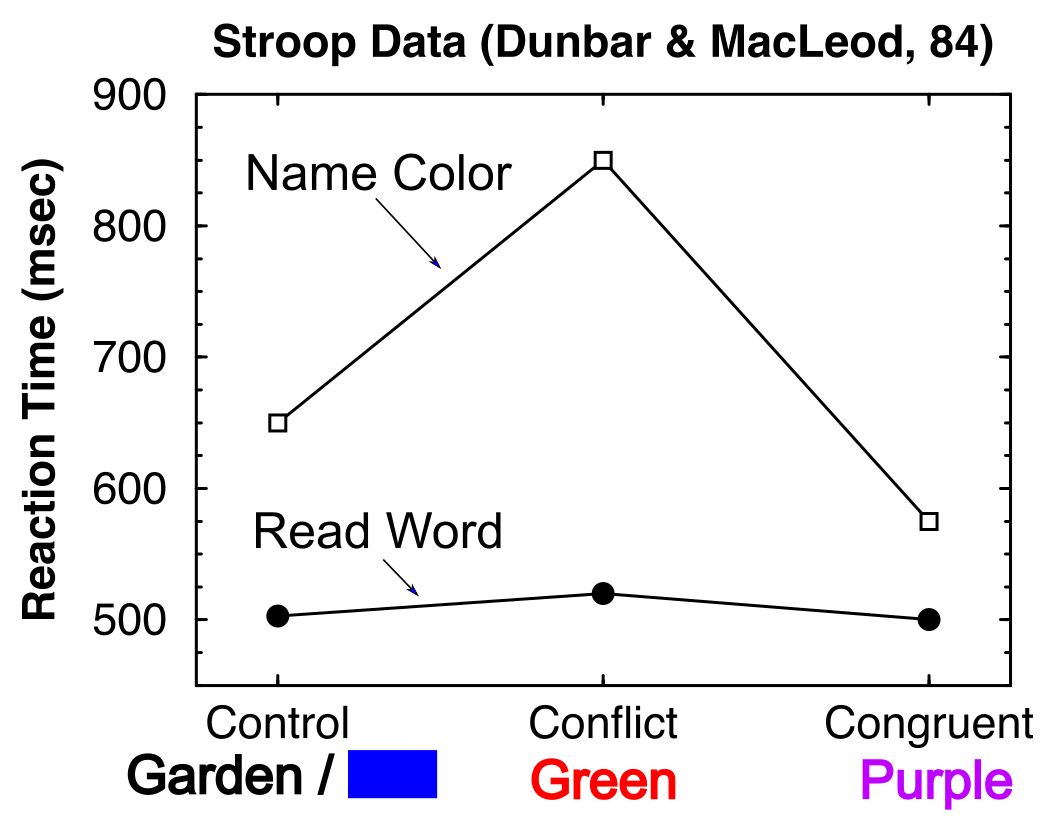
ストループパラダイム ([@fig:fig-stroop]) では、被験者に色の単語 (例: 「赤」、「緑」) が 1 つずつ提示され、その単語 (例: 「赤」) を読むか、その単語が書かれているインクの色の名前を答えることが求められます。 場合によっては、「赤」という単語が 緑 インクで表示され、これは 不一致 または 矛盾 状態を表します。 「ストループ効果」とは、特に色の命名 ([@fig:fig-stroop-data]) の場合、この不一致の条件でエラー率と応答時間が大きくなるというものです。不調和な状態で色の名前を付けるのが特に難しいのは、単語を読むという比較的「自動的」でよく練習された性質が原因であるため、単語を読むという自然な傾向がインクの色に注意を払って名前を付けるのを妨げていると考えられています。
コーエンら。 (1990) Stroop モデルは、維持された PFC タスク表現が、強力な単語読み取り経路に直面して、弱い色処理チャネルをサポートする強力なトップダウン バイアスをどのように提供できるかを示しました。彼らは、単語の読み方と色の名前の違いを、単にこれらの各タスクで提供されたトレーニング量の関数として確立することができました。私たちのシミュレーションは、これらと同じコア機能を再現します。
ストループ モデルは、実行機能における阻害の役割を明確にするのに役立ちます。多くの人は、ストループ タスクについて、インクの色に集中するために強力な単語読み取り経路を抑制する必要があると説明しており、このモデルには抑制力学も含まれています。ただし、このモデルの PFC は、特に単語読み取り経路に指向性の抑制を与えるものではありません。代わりに、弱い経路(色の名前付け)に「興奮性」のトップダウンサポートを提供し、この経路がより支配的な単語読み取り経路と(側方抑制相互作用を介して)よりよく競合できるようにします。したがって、抑制は皮質全体で機能するより付随的な自動プロセスとみなされ、トップダウンのバイアスは無関係な情報を抑制するのではなく、関連する情報を興奮させることに関与しています。同様に、人々は、A-not-B 課題を、以前に成功した行動を繰り返すために強力な反応の抑制が必要であると見なしていましたが、A-not-B モデルや実行機能 [@MortonMunakata02; @StedronSahniMunakata05] を開発する他のモデルは、代わりに、競合する関連するオプションの興奮によって成功します。
トップダウンの認知制御の探求
CCNシムズ で stroop モデルを開きます。
PFC エリア内の下部構造: ストライプ
各機能的 PFC 領域内には、ハイパーカラム、マクロカラム、または ストライプ へのニューロンの興味深い地形的組織化が存在します (これらの用語は通常、同様のタイプの神経組織化に関連付けられていますが、皮質の異なる部分にあり、ストライプは PFC に特有のものです; [@LevittLewisYoshiokaEtAl93])。皮質のすべての領域において、神経トポロジカル構成の最小レベルを 皮質柱 または 微小柱 (より大きな マクロ柱 とより明確に区別するため) として識別できます。これには、直径約 50 ミクロンの領域に約 20 個の錐体ニューロンが含まれています。ストライプにはこれらのマイクロカラムがおよそ 100 個含まれており、通常は幅がおよそ 5 マイクロカラム (250 ミクロン)、長さが 20 マイクロカラム (1000 ミクロンまたは 1 ミリメートル) の細長い形状に組織されています。このような各ストライプは、およそ 10 個以上の他のストライプのセットと相互接続されており、これらを ストライプ クラスター と表すことができます。人間の前頭皮質の大きさを考慮すると、前頭皮質 (運動野を含む) 内には 20,000 もの縞がある可能性があります。
PFC やその他の領域では、マイクロカラム内のニューロンは非常によく似た情報をエンコードする傾向があり、モデルで通常使用される種類の単一のレート コード化ニューロンと同等であると考えることができます。次に、個々のストライプには、そのようなレート コード化されたニューロンに相当するものが約 100 個含まれていると考えることができます。これにより、マイクロカラム全体にわたる疎な分散表現を使用して、かなり多数の異なるものをエンコードするのに十分な余地が提供されます。
機能的には、PBWM モデルでは、BG システムを介して、関連するニューロンのストライプとの接続の対応するストライプごとのループによって各ストライプを独立して更新できるという仮説を立てます。これにより、次に説明するように、BG による PFC での情報の更新と保守に対する非常にきめ細かい制御が可能になります。
大脳基底核とダイナミック ゲーティング
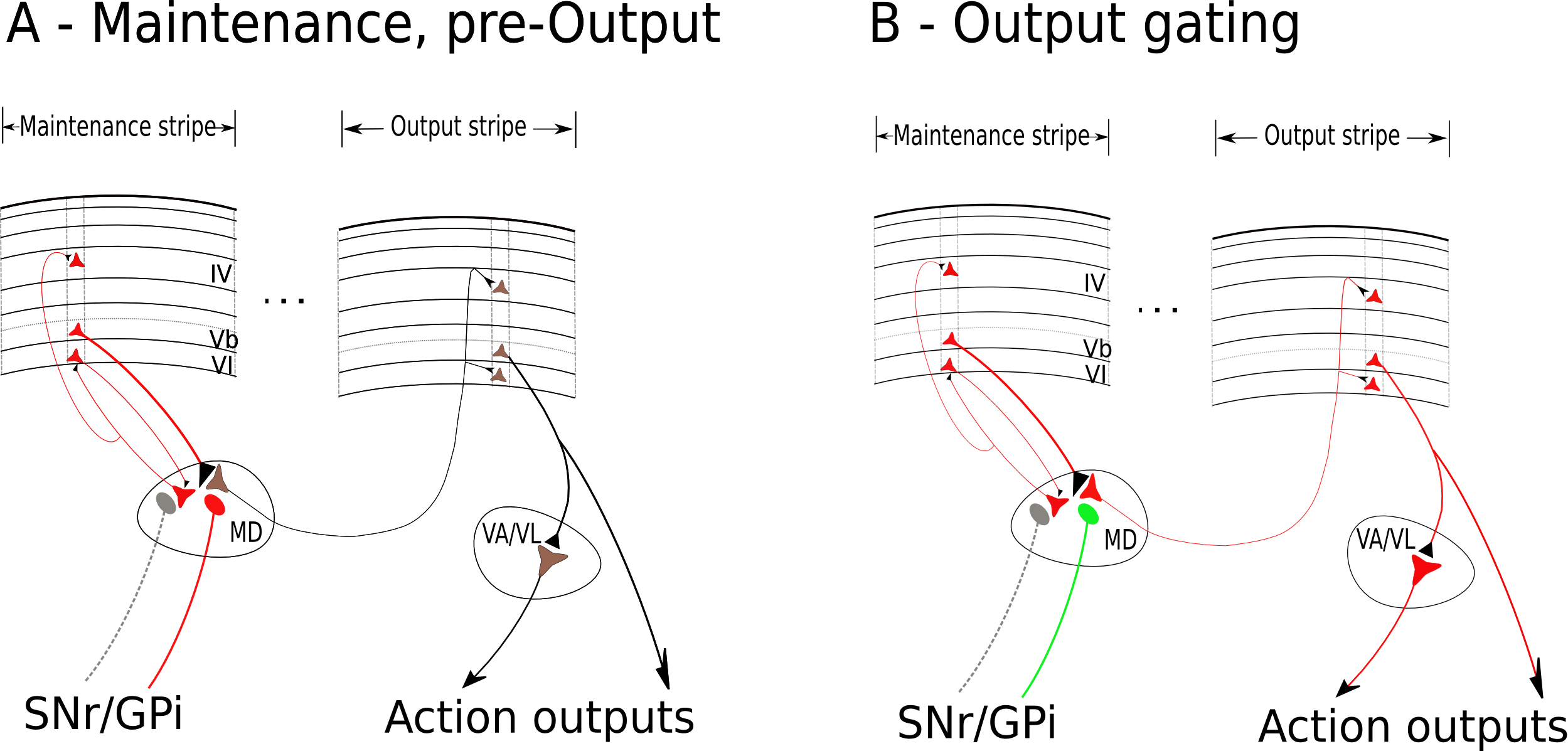
![大脳基底核 (BG) が PFC の能動的維持をどのように調節できるか。 (A) BG 活動がない、または線条体で NoGo (間接的) 経路が発火しているデフォルト状態では、PFC は情報をアクティブな状態に維持し続けます。 SNr (網状黒質) または GPi (淡蒼球内部節) は、視床のニューロンを阻害する強直性 (持続的) 活性を示し、それによって視床皮質ループを停止させます。 (B) Go (直接) 経路の発火は、PFC ストライプの更新をトリガーして、新しい情報 (行動に関連する新しい刺激入力など) をエンコードします。これは、SNr/GPi ニューロンを抑制することで視床皮質ループを開き、PFC の活動のバーストを引き起こし、神経発火の新しいパターンへの更新を促進します。これには、今後もこの新しいパターンを維持し続ける新しい固有の維持電流が含まれます。このメカニズムは、並列回路の前/運動領域との BG 相互作用を介してモーター動作をゲートするメカニズムと同じです (モーターの章を参照)。 [@OReilly06] より転載。](https://raw.githubusercontent.com/compcogneuro/book/main/figures/fig_bg_gating_science.png)
運動制御と強化学習 の章で説明したように、大脳基底核 (BG) は、PFC ([@fig:fig-bg-gating-science]) と双方向に接続されている視床ニューロンの抑制を制御することにより、PFC の活動を調節する立場にあります。線条体活動がない、または間接 (NoGo) 経路ニューロンの発火というデフォルト状態では、SNr (黒質網様体) または GPi (淡蒼球内節) ニューロンが視床を強張的に抑制します。これにより、視床皮質ループの活性化が妨げられ、運動動作の開始や PFC のアクティブな記憶表現の更新に重要であると考えられているのは、このループの活性化です。線条体の Go (直接) 経路ニューロンが発火すると、強直性 SNr/GPi 阻害が抑制され、それによって興奮性視床皮質ループの活性化が可能になります。この興奮性活性化の波は、PFC ニューロンの新しい集団を活性化することができ、その後、新しい Go 信号が発火されるまで積極的に維持されます。
フェーズ的 DA と一時的なクレジットの割り当て
運動制御と強化学習の章でも説明した、実行機能のもう 1 つの重要な生物学的メカニズムは、中脳 (腹側被蓋野 (VTA) および 黒質緻密部 (SNc)) における相性ドーパミン (DA) ニューロンの発火です。これらのニューロンは、最初は一次報酬(リンゴジュースなど)に反応しますが、その後、これらの一次報酬を確実に予測する条件刺激(CS)の開始時に発火するように学習します。これは、PFC アクティブ メンテナンス システムの重大な問題を解決する タイム トラベル の一種に相当します。つまり、何をメンテナンスするかについての決定は、通常、何か有用なものをメンテナンスしたというその後の価値よりもかなり前に行われることを考えると、システムは何をメンテナンスするかをどのように学習するのかということです。 PFC での有益な情報の維持を一種の CS と考えると (確実にポジティブな結果と関連付けられるはずなので)、ドーパミン ニューロンはそのような CS の開始時に発火することを学習します。 CS の開始時にこの位相的 DA 信号が得られると、そもそもこの情報を維持するという決定が強化され、タイムトラベルの問題が解決されます。
次に説明する計算モデルには、CS 開始時の位相 DA 信号によって、新しい情報を PFC アクティブ メンテナンスに更新する BG Go ニューロンの学習を駆動させることにより、この重要なアイデアが組み込まれています。このモデルは、この中心となるアイデアが複雑な実行機能タスクの学習をサポートするのに十分であることを示しています。
PBWM 計算モデル
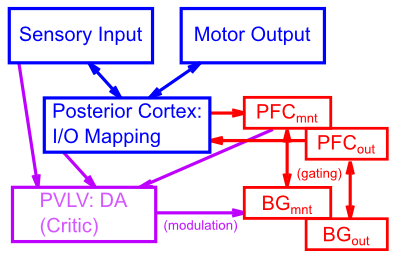
上で検討した PFC/BG システムの生物学的特性は、PBWM (前頭前皮質大脳基底核ワーキング メモリ) [@OReillyFrank06; @HazyFrankOReilly06; @HazyFrankOReilly07] ([@fig:fig-pbwm-architecture-mnt-out]) と呼ばれる計算モデルで捕捉されます。このモデルの PFC ニューロンは、個別に更新可能なストライプに編成され、また、メンテナンスと出力ゲートの個別の機能グループにも編成されます (詳細は後述)。さらに、各 PFC ストライプは、表層 (2、3) と深層 (5、6) の観点から表されます。深層ニューロンは、特に、NMDA および mGluR チャネルと視床を通る興奮性ループの効果を表す、さまざまなメカニズムを通じて長時間発火を維持する能力を持っています。特定の PFC ストライプの表層から深層へのアクティベーションの流れは BG ゲート信号に依存し、BG 層も対応するメンテナンス ストライプと出力ゲート ストライプに編成されます。 BG の Matrix 層 (線条体のマトリソームを表す) には、GPi と視床の組み合わせ (GPiThal) 層に投影する別々の Go ニューロンと NoGo ニューロンがあり、ストライプごとに 1 つのニューロンがあり、Go 経路が NoGo よりも十分に強い場合に発火します (このメカニズムは、GPe、GPi/SNr、STN、および視床を含む詳細な BG ゲート回路を抽象化しています)。モーターの章でシミュレートされているように、機能は単一の GPiThal レイヤーに単純にまとめられています)。 GPiThal Go 信号は、現在の表層レイヤーのアクティベーションを反映するように PFC 深層アクティベーションを更新しますが、NoGo は PFC をそのままにして以前の情報を維持し続けます (またはまったく何も保持しません)。
PVLV 相性ドーパミン システムは、運動制御と強化学習 の章で説明されているのと同じ強化学習メカニズムを使用して、BG Go および NoGo ニューロンの学習を促進します。正の DA バーストは Go の促進と NoGo 重みの抑制につながり、DA ディップではその逆になります。
おそらく、PBWM システムがどのように機能するかを理解するための最も重要な鍵は、BG でさまざまなゲート戦略を 試行錯誤 して探索し、DA がプラスの報酬に関連する戦略を強化し、そうでない戦略を罰するということです。モデルの現在のバージョンでは、マトリックス学習は報酬時のドーパミン発火によってもっぱら駆動され、シナプスタグベースのトレースメカニズムを使用して、このドーパミン作動性の結果につながった以前のすべてのゲートアクションを強化/罰します。具体的には、特定のマトリックス ユニットがゲート動作のために発火すると、アクティブな入力を持つシナプスが シナプス タグを確立します。このタグは、後続の相性ドーパミン作動性結果信号まで持続します。広範な研究により、シナプス内のアクチン線維ネットワークに基づくこれらのシナプスタグは最長 90 分間持続することができ、その後強力な学習イベントが発生すると、タグ付けされたシナプスも [@RedondoMorris11; @Rudy15; @BoschHayashi12] によって強力に強化されることが示されています。この形式のトレースベースの学習は、以前のゲーティング イベントの報酬の影響について学習するために他のメカニズムを必要としないため、計算的に非常に効果的です。 PBWM モデルの以前のバージョンでは、ゲーティングを強化するために CS (条件刺激) ベースの相性ドーパミンに依存していましたが、このスキームでは、PFC が維持する活性化が一種の内部 CS 信号として機能すること、および扁桃体がこれらの PFC 活性化状態をデコードして有用なアイテムがメモリにゲートされたかどうかを判断することを学習する必要があります。トレースベースのメカニズムと比較して、この CS ドーパミンのアプローチははるかに複雑でエラーが発生しやすくなります。代わりに、一般に、マトリックス学習を推進する CS は標準的な外部タイプであり、望ましい結果に向けての進歩を知らせ、その中間状態に至るまでのアクションを強化すると仮定します (つまり、CS は サブゴール の達成を表します)。
通常、複数のストライプの存在は、PBWM モデルが迅速に学習するために重要です。複数のストライプの存在により、単一のストライプがすべての異なる戦略を順番に探索するのではなく、異なるゲート戦略を 並列 で探索できるためです。 1 つのストライプが有用なゲート戦略をヒットできる限り、システムは成功し、他のストライプを無視してその有用なストライプに焦点を当てる方法をすぐに学習します。複数のストライプは、タスクの進行中に複数の情報を維持および更新する必要がある場合にも重要です。実際、この需要こそが、初期のゲーティング モデルに取って代わる元の PBWM モデルの開発の動機となったのです。このモデルでは、位相ドーパミン信号を使用して PFC 表現を直接ゲート制御していましたが、マルチ ゲートはサポートされていなかったため、単一アイテムの容量に制限されていました。これらの複数のストライプがあることの興味深い結果の 1 つは、「迷信的な」ゲートが他のストライプでも発生する可能性があることです。そのゲートが実際に役立つゲート信号と十分に確実に一致する場合、それも強化されます。おそらく、これは私たちの迷信深い傾向を明らかにするかもしれませんか?
出力ゲート
[@fig:fig-SommerWurtz00-fig2] で見たように、一部の PFC ニューロンは遅延期間 (アクティブ メンテナンス) 発火を示しますが、他の PFC ニューロンは出力応答発火を示します。これらの集団は混合しているようには見えません。通常、特定のニューロンが両方のタイプの発火の組み合わせを示すことはありません。これは、メンテナンス ゲート ではなく 出力ゲート される PFC ストライプの別のセットを持つことによって PBWM フレームワークで捕捉されます。つまり、維持された情報は、下流の処理 (注意やモーター応答の選択など) に影響を与えるべきかどうかを決定するために、さらなるゲートの対象となる可能性があります。通常、メンテナンスと出力ゲート ストライプの単純なペアを使用し、メンテナンスから出力 PFC ユニットへの直接 1 対 1 の投影を使用しますが、これらのストライプ間には任意の形式の関係が存在する可能性があります。ただし、出力 PFC ユニットは、対応するストライプ レベルの BG/GPiThal Go 経路が起動した場合にのみアクティブになります。したがって、情報は、行動を促進するためにアクティブに出力される前に、アクティブではあるがある程度「オフライン」の形式で維持できます。 [@fig:fig-mnt2out] は、メンテナンス側と出力側の間でのゲーティングの分業と、「ハンドオフ」がどのように発生するかを示しています。
出力ゲーティングに関するさらなる考慮事項、(出力ゲーティング後) 不要になったときに保持されている情報が消去される方法、学習の改善に役立つゲーティング バイアスなど、PBWM の詳細については、関連する方程式とデフォルト パラメーターも含まれている PBWM の詳細 付録を参照してください。
PFC アクティブ メモリの動的更新: SIR モデル
Stroop モデルと A-not-B モデルで、持続的な PFC 活動がトップダウン バイアスを通じてどのように行動に影響を与えるかを確認しました。次に、大脳基底核による PFC 表現の動的なゲート制御や、情報を迅速に更新して堅牢に維持する機能など、PFC 機能のより複雑な側面に目を向けます。 PBWM モデルによってキャプチャされたこの機能の最初の導入として、単純な SIR (Store、Ignore、Recall) タスクを使用します。このタスクのトライアルのサンプル シーケンスを次に示します。
-
S - A — これは、ネットワークが後で呼び出せるように A 刺激を保存する必要があることを意味します — ネットワークは A に応答します。
-
I - C — C 刺激を無視しますが、それでも応答する必要があります — ネットワークは C に応答します。
-
I - B — B 刺激を無視します — ネットワークは B に応答します。
-
R — 最後に保存された刺激を呼び出します — ネットワークは A に応答します。
BG メンテナンス ゲート システムは、後で呼び出すために関連する刺激をエンコードするために、ストア トライアルで PFC の更新を駆動するために Go を起動することを学習する必要があります。また、以前に保存された情報が上書きされないように、無視刺激に対して NoGo を起動する方法も学習する必要があります。最後に、リコール トライアルでは、出力 BG ゲート メカニズムが PFC からの保存された情報の出力を駆動する必要があります。ネットワークは、これらのさまざまな入力のセマンティクスについて何も知らない状態で開始され、さまざまな入力をどう処理するかを完全に試行錯誤を通じて学習する必要があることを理解することが重要です。
PFC アクティブ メモリの動的更新の探求
この学習の展開を確認するには、CCNシムズ で sir モデルを開きます。
簡略化のためにここでは考慮しませんが、この PBWM モデルの真の力は、複数の並列ストライプを持つように拡張されたときに発揮されます。したがって、作業記憶内の複数の情報を個別に更新および維持すること、および必要に応じて正しい情報を取得することを学習できます。この要求の良い例は、SIR-2 タスクに要約されています。このタスクでは、単一のストアおよびリコール タスク制御信号が関与する代わりに、2 つのそのような信号 (つまり、S1 と S2、および R1 と R2) が存在します。したがって、ネットワークは 2 つの刺激を別々に保存し、それらを別々のバッファーに更新し、R1 と R2 を呼び出す合図のときに正しいバッファーに保持されている情報に基づいて適切に応答することを学習する必要があります。このモデルのより最近の変形は、連続的な刺激 (色など) の作業記憶に適用され、モデルが記憶表現の精度の変化を考慮できるようになりました (Soni & Frank、2024; https://elifesciences.org/reviewed-preprints/97894 を参照)。このモデルはさらに、複数のメモリ表現を「チャンク」にマージできる適応型ゲーティング戦略を学習し、パフォーマンスを向上させます (多数のストライプを備えたモデルを超えてさえ)。これは人間のパフォーマンスと一致しており、メモリ内で複数の項目を管理する方法を学習する難しさに基づく作業記憶容量の制限の説明になりますが、チャンク化によって軽減されます。
PFC アクティブ メモリのより複雑な動的更新: N-Back タスク
N バック タスクは、機能的 MRI (fMRI) 実験において PFC の活性化を非常に確実に推進できるため、PFC における複雑な作業記憶機能の最も広く使用されている尺度の 1 つとなっています。 Chatham ら (出版中) は、このタスクの PBWM ベースのモデルを開発しました。これは、より複雑な認知タスクが PBWM によってどのように学習できるかを示しています。このモデルは、認知神経科学の文献におけるさまざまな重要な発見にも触れています。
PFC の階層構造: サブタスク、目標、認知的順序付け
![複数の前頭前大脳基底核ループにわたる階層的なアクションの選択。一番右の最前レベルでは、PFC は、下位レベルのアクションをガイドするためにこのコンテキストを維持することが報酬を予測する確率に基づいて、対応する BG ループによってゲートされるコンテキスト情報を表します。中央のループには、入力ゲートと出力ゲートの両方が含まれます。入力ゲート メカニズムにより、刺激表現 S が PFC_maint 層を更新できるようになりますが、出力ゲート メカニズムは、前方 PFC のコンテキストを条件として、維持された情報のサブセットをゲート アウトします。関連する BG 層は、維持された刺激 S とコンテキストを考慮して、出力ゲーティングの報酬確率を学習します。最後に、一番左のモーター ループは、モーターの章で説明した単一ループ BG モデルと同様に、刺激を条件とした報酬確率に基づいて単純なモーター応答をゲートすることを学習しますが、ここでは関連する刺激の特徴がより前方のループによって選択されます。 [@BadreFrank12] から転載。](https://raw.githubusercontent.com/compcogneuro/book/main/figures/fig_frank_badre12_pfc_hierarch.png)
複数の PFC-BG 回路にわたるアクションの階層制御をシミュレートする関連モデルについては、[@ReynoldsOReilly09; @FrankBadre12; @CollinsFrank13] を参照してください。後者のモデルでは、特定の感覚状態でどのアクションを選択するかを示す潜在的なルール セットが複数存在し、適切なルール セットがより高いレベルのコンテキストに依存する可能性がある状況を考慮します。 (たとえば、誰かにハグ、キス、握手、手を振るなどの挨拶をする傾向は、その人との関係、街にいるのか職場にいるのかなど、状況によって異なる場合があります。また、新しい国 (または都市) に行く場合、適用するルール セットが他の国で適用したルール セットと同じである場合もあれば、新しいルール セットの作成が必要になる場合もあります。)より一般的には、より高いレベルのルールを、さまざまな刺激に応じてどのように行動するかを文脈化した「タスクセット」と呼びます。階層型 PFC-BG ネットワークは、これらの PFC タスク セットの作成方法を学習し、同時に各タスク セットでどのアクションを選択するかを学習できます ([@fig:fig-frank-badre12-pfc-hierarch])。
重要なのは、この階層表現を使用すると、学習された PFC 表現が抽象的であり、その表現を指示するコンテキストから独立しているため、一般化と他のコンテキストへの転送が容易になると同時に、新しいタスク セットを作成する必要がある時期も特定できることです。また、既存の抽象的なタスク構造に新しい知識を追加することもでき、その知識は、それを手がかりとする他のコンテキストにすぐに転送できます (言語で新しい単語を学習するのと同じように、その単語をすぐに他のコンテキストや他の人々と再利用できます)。一般化と転送のデモンストレーションを含め、このネットワークの動作を確認するには、ここ にリンクされている Collins & Frank ネットワークを参照してください。このモデルをテストしたさまざまな経験的データは、実際に人間 (赤ちゃんも含む!) が、一般化と転移を容易にするような方法で、そのようなタスクセットを階層的な方法で (そうするように指示されていない場合や、学習に有益でない場合でも) 表現していることを示しています。そして、この階層構造の範囲は、PFC および BG [@BadreFrank12; @CollinsFrank16a] の神経署名に関連していると考えられます。
上記で検討した要素の多くを最も重要な用途に活用するために、BG ゲーティングと併せて、PFC のさまざまな領域 (以前に検討した感情領域を含む) の調整された相互作用によって、システムが認知処理の複数の連続ステップにわたって一貫したタスク主導型の方法で動作できるようにする方法を検討します。これはまさに人間の知性の特徴です。柔軟で適応的な方法で、一連の単純な認知ステップを実行することで、複雑な問題を解決できます。 ACT-R などのより抽象的な認知モデルは、このレベルの認知の機能的特性の優れた特徴付けを提供します。ここで検討するモデルの目的は、より詳細な神経メカニズムがどのように連携してこの機能を生み出すことができるかを理解することです。
-
より高い(より前方の)レベルの PFC は、一連の認知行動を組織化するためのコンテキスト/目標/計画をエンコードし、より低い、より後方の PFC 領域によって駆動されます。重要なことに、これらの上位領域は、アクションの厳密なシーケンスを指定するのではなく、むしろアクションのシーケンスの望ましい結果状態をコード化し、適切な下位レベルのステップが選択されるように適切なコンテキストを提供します。
-
一連のアクションの各ステップには、他の可能なオプションと比較したアクションの報酬結果と労力コストの考慮が含まれます。
執行機能に対する感情的な影響: OFC と ACC の役割
PFC と実行機能の最も重要な特徴の 1 つは、感情的および動機付けの影響を高度な認知制御と計画とともに統合していることです。 PFCの内側領域と腹側領域は感情的要因や動機付け要因の処理に特に重要であり、眼窩前頭前皮質(OFC)を含む腹側内側領域は刺激の感情値をコード化するのに重要であり、背側内側領域(主に前帯状皮質(ACC))は運動行動や計画の感情値をコード化するのに重要であるという考えにかなりのデータが集まっている。
その他の実行機能
上記で検討した主なモデルは、実行機能の最も中心的かつ重要な側面のいくつかをカバーすることを目的としていますが、これは非常に広い領域であり、残念ながらカバーできない重要な現象が多数あります (ただし、将来の改訂ではカバーされる範囲を拡大し、オプションのモデルで以下のさまざまなトピックをカバーする予定です)。
多くの人、特に学術的な環境にいる人にとって、高次の認知機能に名前を付けるように求められた場合、最初に思い浮かぶのは次のようなものかもしれません。あるいは、おそらく、重要な決定を下すために慎重な論理的推論を使用することもあります。しかし、これらの高度に形式化された領域に加えて、非常に洗練されたレベルの処理を伴う、日々の、しかしそれでも重要な精神的活動が数多くあります。たとえば、次のような活動です。水着の季節までに10ポンド減量しようとしているときにデザートを食べたいという誘惑に抵抗する、または;ブラックジャックをプレイしながら作業記憶でカードを数えます。これらすべての種類の精神活動は、その最適な表現のために前頭葉皮質と関連構造に依存していることが現在では知られています。以下は、特徴的な実行機能のいくつかの主要なカテゴリのリストです。
-
多くの場合、形式記号システムが関与する、高度に構造化された認知活動 — 数学、形式論理、コンピューター プログラミング、創造的および/またはノンフィクションの執筆、および構造化された合理的な意思決定の学習および/または使用などの精神的活動。これらはすべて、タスク関連情報、特に高度に抽象的で象徴的な性質の情報を時間的に延長して維持する必要があります。これらおよび他の多くの実行機能における言語の役割は非常に重要な側面です。言語は、行動の積極的な維持と制御のための非常に柔軟な精神通貨を提供します。特定の単語やフレーズを思い出すことで、達成したいことや最初の処理ステップで何を導き出したのかなどを思い出すことができます。
-
海馬におけるエピソード情報のエンコードと検索の制御 — 海馬と PFC/BG システムは、さまざまな形態の実行機能において大きく相互作用しており、海馬の急速な学習能力が PFC の一時的で柔軟な能動的維持特性を補完している可能性が非常に高いです。 PFC が気を散らされると、通常、情報はアクティブなメンテナンスから永久に失われてしまいますが、海馬は長期にわたるシナプスの変化という観点から情報を符号化し、取得することができます。多くの場合、PFC で情報を永続的にアクティブに維持するよりも、この海馬のエンコーディングと検索を使用する方が効率的である可能性があります。
代替フレームワークとモデリングアプローチ
この章では、1 つの特定の理論的枠組みに焦点を当てましたが、他にも多くのアプローチが長年にわたって説明されてきました。おそらく最も影響力のあるモデルは [@Baddeley86] によるもので、彼は特に作業記憶に焦点を当てましたが、同時に「中央幹部」についても主張しました。特に、彼は作業記憶の 2 つの特定の形式を仮定しました。1) 言語情報を維持するための 音韻ループ。 2) 空間情報用の 視覚空間スクラッチパッド。もう 1 つの非常に影響力のある理論的アプローチは、監督注意システム (SAS) フレームワークについて説明した [@Shallice88] によるものです。最後に、非常に影響力のある従来の AI アプローチもあります。これについては、以下で簡単に説明します。
従来の AI は主に上記に列挙した種類の認知機能によって動機付けられ、特定の種類の問題を解決するために何が必要かを解明し、そのように物事を行うモデルを設計することに重点を置いた、シンボルを使用した設計指向のアプローチに主に焦点を当ててきました。このアプローチには皮肉があり、このアプローチを採用している研究者は、説明しようとしている非常に高いレベルの認知機能を、それを再現するシステムを設計するために使用していることになります。この種のアプローチの根本的な問題は、基本的に説明しようとしている機能そのものを設計していることです。これは、この種のアプローチにメリットがないと言っているのではなく、最終的に説明できる内容が限られているというだけです。おそらく明白な理由から、これらの種類の認知機能モデルは、最高レベルとして列挙した種類の認知機能、つまり形式的な数学と論理を実行できるモデル化システムを扱う場合に最も成功しています。彼らがあまりうまくやってこなかったのは、あまり高レベルではないと考えられたり、さらには低レベルであったり、私たちが自動的に行うものだとみなしがちなことの多くを説明することです。生物学的に情報を与えられたニューラル ネットワークのアプローチが最も役立つのは、後者の領域です。したがって、これら 2 つのアプローチはうまく補完することができ、ハイブリッド アプローチが追求されています。たとえば、Leabra アプローチは、SAL と呼ばれるアーキテクチャで ACT-R アプローチとハイブリッド化されています。
これらのアプローチはすべて相互に排他的ではなく、多くの共通のアイデアを共有しており、多くの点で補完することができます。特に、従来の AI アプローチは、高レベルの問題 (算術など) の解決に直接取り組むことで、最も高度な認知機能の一部に対処できます。一方、私たちが提唱するニューラル ネットワーク アプローチの目標は、基礎となる自動メカニズムに基づく制御のような処理の出現に対する還元主義的な説明を提供しようとする、よりボトムアップのモデルを提供することです。これが PBWM フレームワークで採用されるアプローチです。
重要なポイントのまとめ
-
前頭前皮質 (PFC) は、持続的な神経発火を通じて活動状態にある情報をエンコードします。これは、シナプス重みの変化を使用するよりも柔軟で、迅速に更新可能です。
-
大脳基底核 (BG) は、PFC のアクティブな記憶状態の更新 (動的ゲーティング) を駆動し、柔軟性を高めます。
-
中脳核からの位相性ドーパミン信号は、その後の報酬を予測する刺激の開始よりも早い時期に報酬の関連付けを転送することにより、BG ゲーティングのトレーニングに適した特性を持っています。
-
PFC は、トップダウンの興奮性バイアスを介して脳内の他の場所の認知処理に影響を与えます。 A-not-B モデルは、PFC の積極的な維持能力の出現が、乳児が発達の過程で示す行動の変化をどのようにサポートできるかを示しています。
*ストループ モデルは、タスク情報のアクティブなメンテナンスがどのように行動を導き、矛盾するオプションを克服するのに役立つかを示しています。
-
BGダイナミックゲーティングは、一部の情報を動的にエンコードし、その他の無関係な情報を無視し、アクティブメモリの内容を更新することで、柔軟な認知機能をサポートできます。 SIR モデルと n-back モデルは、これらの機能を実証します。
-
PFCの内側領域と腹側領域(眼窩前頭前野(OFC)と前帯状皮質(ACC))は、それぞれ刺激と行動に関する感情情報を伝え、取るべき潜在的な行動(意思決定、問題解決など)を適切に評価するために重要です。