compcogneuro/web: cerebellum
このページは外部資料の日本語訳です。原文の見出し順と本文順を保ち、コード・URL・出典表記はできるだけ原形のまま残しています。
出典とライセンス
原典: https://github.com/compcogneuro/web/blob/main/content/cerebellum.md
ライセンス: Text: CC BY 4.0; code: BSD 3-Clause。このページは日本語翻訳であり、変更点は翻訳とサイト内整形です。
+++ Categories = [“Neuroscience”, “Cognition”] bibfile = “ccnlab.json” +++ 小脳は、高度に特殊化された神経要素のシステムであり、その多くは脳の他の部分に見られるものとは異なります。この特殊な機構すべての正確な機能は、多くの並行チャネルにわたって複製され、脳内のほとんどの感覚経路および運動経路と接続されていますが、そのユニークな機能の多くに関する独創的な出版物 ([[@EcclesItoSzentagothai67]]) 以来、興味をそそられるほどとらえどころがありません。
小脳は運動制御システムであると広く考えられていますが、小脳は運動システムと密接に相互接続されているものの、解剖学的には感覚システムです。それが学習に関与していることは明らかですが、正確に何をどのように学習するかについては、依然として活発な議論が行われています。
適応フィルター モデル ([[@Fujita82a]]; [[@DeanPorrillEkerotEtAl10]]; [[@Cullen23]]) の特定の実装を採用します。これは、自分自身の運動動作によって生成される予測された感覚信号 (つまり、感覚再フェランス) をフィルターで除去し、運動動作のより安定した有用な基盤を提供します。これらの感覚予測は、順モデル ([[@MiallWolpert96]]; [[@Bastian06]]; [[@ShadmehrSmithKrakauer10]]; [[@TanakaIshikawaLeeEtAl20]]) としても使用され、運動動作の感覚的影響を予測して、より調整されたさらなる動作を推進します。これらは、[[@^Marr69]]、[[@^Albus71]]、および [[@^Ito72]] によって元々提案され、依然として最も広く議論されている ([[@RaymondMedina18]]; [[@Shadmehr20]]) によって最初に提案された、より一般的な [[error-driven learning]] モデルと一致していますが、それよりもはるかに具体的です。
{id=”figure_motor” style=”高さ:25em”}
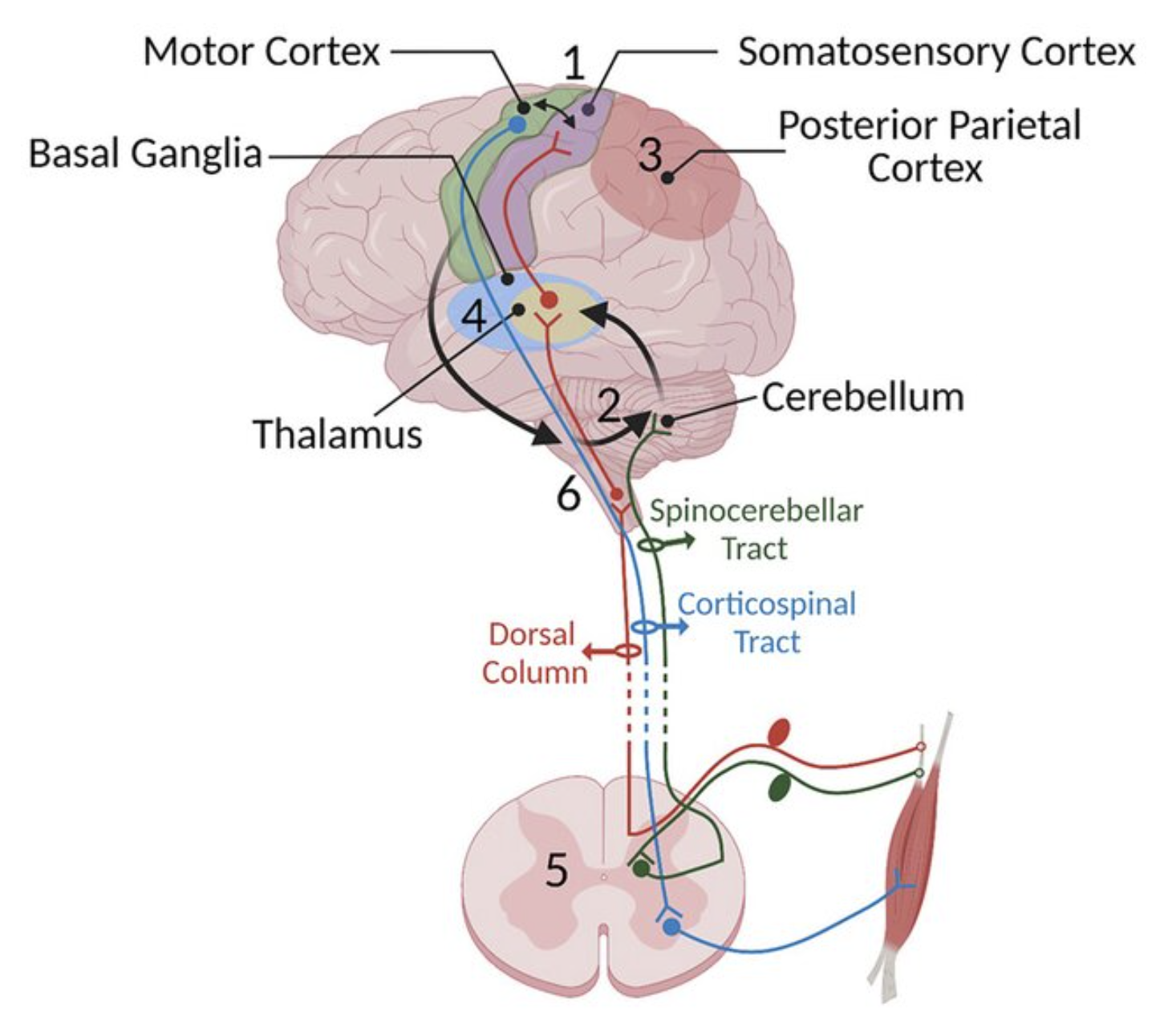
小脳は脳の基部に位置し、中脳の一種の増殖物を形成しています ([[#figure_motor]])。これは、脊髄および中脳の感覚運動野、[[basal ganglia]]、および橋または橋核を介して、一次運動皮質野および体性感覚皮質野(M1およびS1)を含む運動関連皮質野を含む、脳の主要な[[motor]]関連領域のすべてから入力を受け取ります。また、[[vestibular]] システムからの主要な投影を含む、他の幅広い感覚信号も受信します。
深部小脳核 (DCN または最近の用語では単に CN) からの小脳出力は、より外側の CN 領域から VL [[thalamus]] への主要な駆動投影を含め、脳の多くの領域に広く投影され、一次運動野にトレーニング信号を提供できます。小脳は初期学習に重要であるが、その後、皮質を含む他の構造を訓練することによってこの学習を伝達できる可能性は、発生初期の小脳病変の重症度の違いによって裏付けられています([[@BeuriatCristoforiRichardEtAl20]])。
{id=”figure_outer_prod” style=”高さ:15em”}
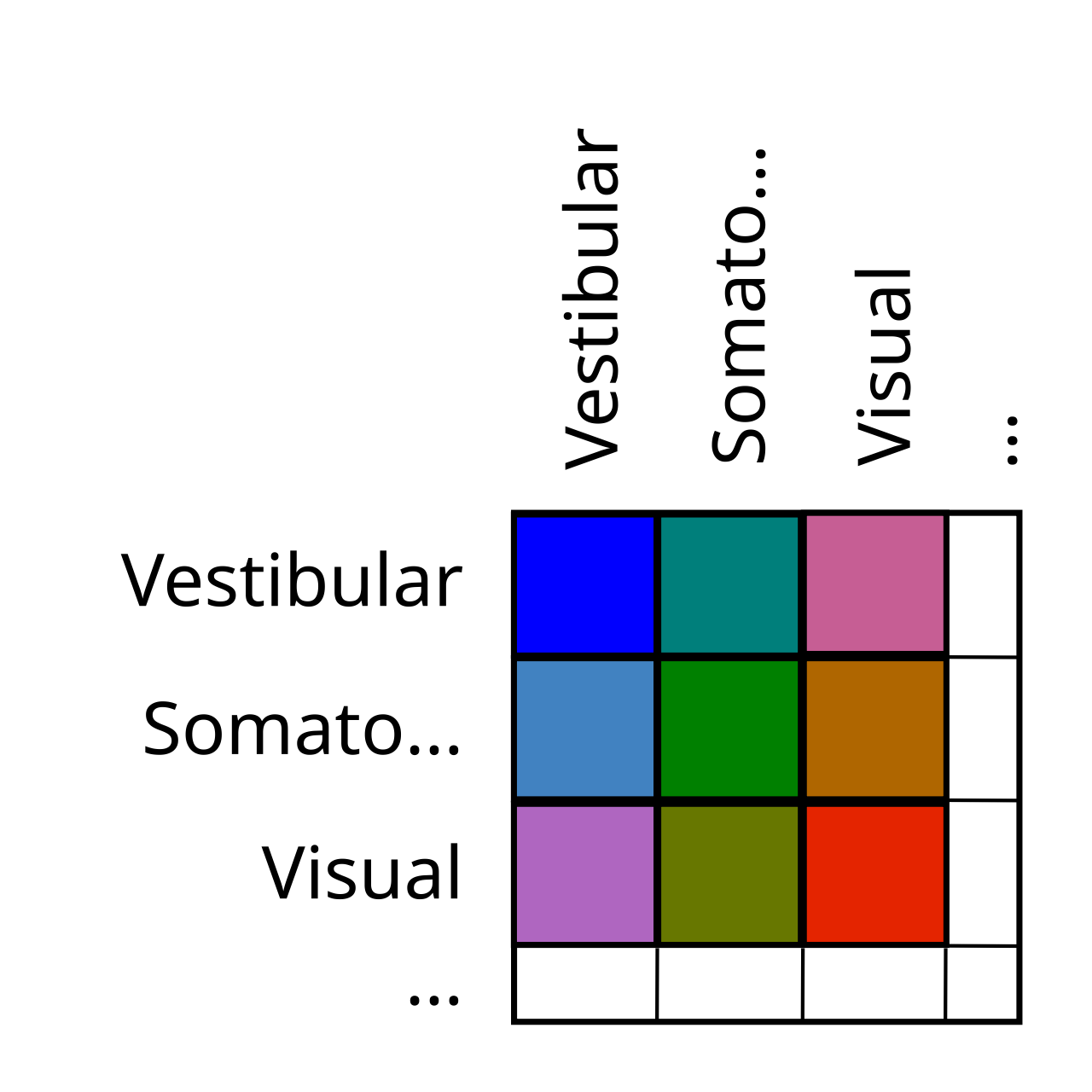
大脳基底核、新皮質、視床とは異なり、小脳は図で簡単にラベル付けできる単純な地形構造を持っていません。代わりに、小脳の主な機能は、さまざまなモダリティにわたる信号を統合して比較することであるため、小脳はさまざまな入力にわたる「外積」のように組織されており、各領域があらゆる組み合わせで 1 つの感覚入力を別の感覚入力と比較します ([[#figure_outer_prod]])。
{id=”figure_fastigial” style=”高さ:45em”}
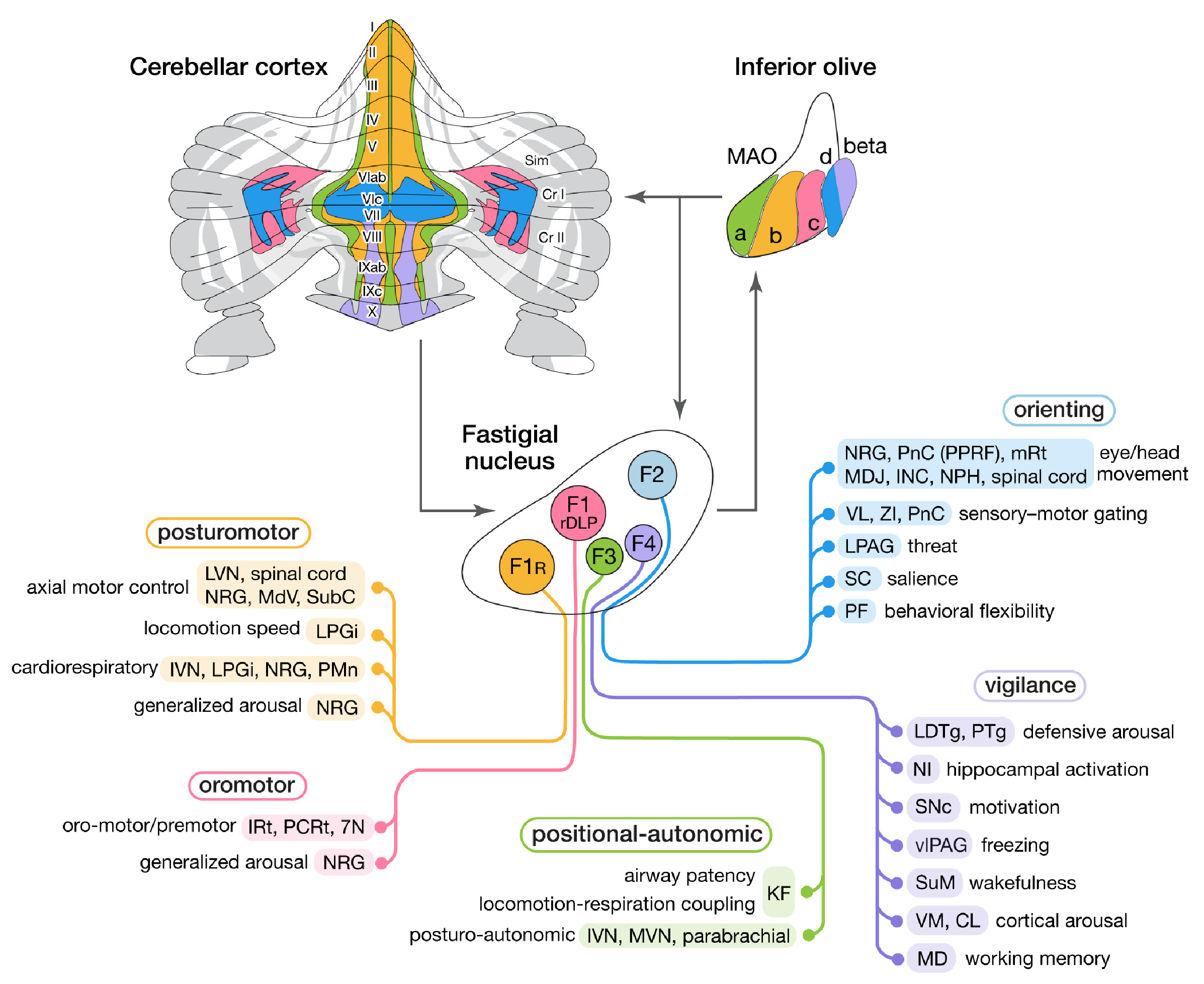
[[#figure_fastigial]] は、進化的に最も古い小脳核 ([[@ZhangWangZhu16]]) である fastigial 核 (FN) に関連する、この外積様組織 ([[@FujitaKodamaduLac20]]) の例を示しています。 FN は主に 小脳皮質の内側_虫_領域と相互接続されており、[[vestibular]]、さまざまな中核生存関連システムに関連する体性感覚および内受容信号などのさまざまな信号が統合されています。
{id=”figure_mossy-prjn” style=”高さ:20em”}

小脳における感覚信号の完全な混合の別の表現は [[#figure_mossy-prjn]] に示されており、橋のニューロンからの単一 苔状線維の軸索の末端領域をプロットしています ([[@BiswasLuoSarpongEtAl19]])。この 1 つの感覚信号は小脳皮質の多くの領域に分布しており、そのようなニューロンの多くは小脳核にも同様に投射します。成体マウスの小脳 ([[@SurchevNazwarWeisheitEtAl07]]) には合計約 4,300 万個のニューロンがあり、これは外積組織の組み合わせの巨大な爆発と、小脳皮質の多数の顆粒細胞による大きな追加乗数を反映しています。
{id=”figure_cortex” style=”高さ:25em”}
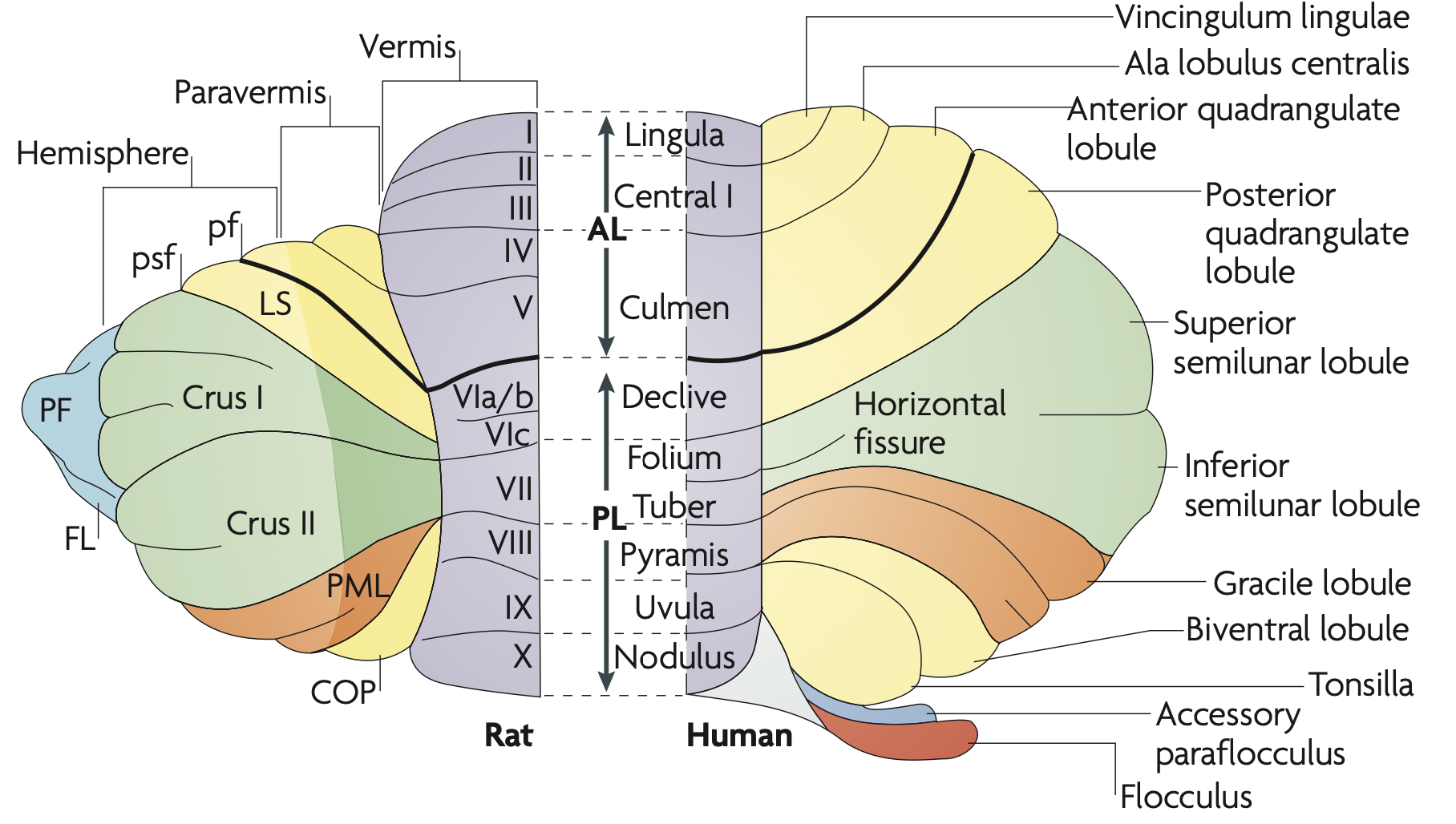
[[#figure_cortex]] は、小脳皮質のさまざまな領域すべての用語を示しています。内側-外側軸に沿って、元の虫部から傍虫部および外側半球部に向かって外側に進み、よりきめ細かい骨格運動制御に関与します。また、前庭信号、視覚信号、および視覚運動信号の収束を処理する_flocculus_ (FL) および paraflocculus (PF) として知られる独特の領域もあります。
[[#figure_mossy-prjn]] に示される縞状の構造は、小脳の長さに沿った長い縞の組織化構造を反映するゼブリン II_ (つまり、アルドラーゼ C または ALDOC) と呼ばれるタンパク質による部分があります ([[@SugiharaShinoda04]]、[[@HawkesLeclerc87]])。小脳の平らなバージョンは視覚化に便利ですが、実際には大規模に折り畳まれた構造であり、折り目はゼブリン縞の長軸に対して垂直になっています。
{id=”figure_cortex-nuclei” style=”高さ:30em”}

[[#figure_cortex-nuclei]] は、小脳皮質と、下オリーブ核 (IO) のさまざまな核および領域との関係を示します ([[@AppsHawkes09]] より)。核の名前にはさまざまな用語があり、齧歯動物の名前は、内側 (つまり、側核) から_側方_、その間に_中間核、または_中間核_と_間質_核が入ります。外側核は進化的に最も現代的な核(霊長類では「歯状」と呼ばれる)であり、ヒトで最も拡大した領域である外側半球と相互接続しています。高度に器用な手の動きに伴う細かな運動制御がサポートされています。中間核は、陸上に住む動物で進化した四肢の主要な骨格筋制御に関与しています。
{id=”figure_somatotopy” style=”高さ:15em”}
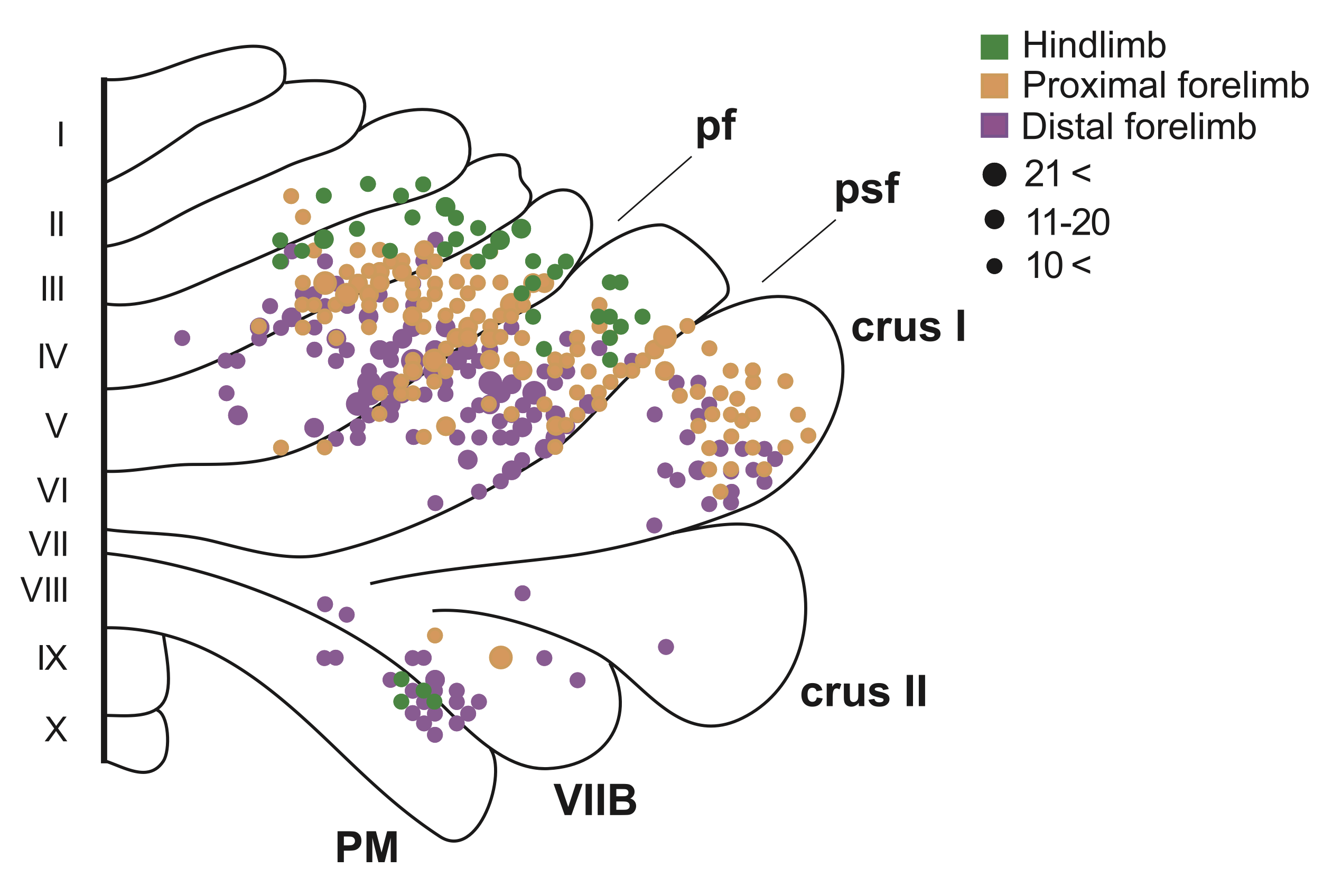
[[#figure_somatotopy]] は、介在/間位 (IP) 核と相互接続されている骨格筋と相互接続されている小脳の部分の体性 (地形的) 組織化の例を示しています。一次体性感覚皮質 (S1) と運動皮質 (M1) が完全に体性局所的な「ホムンクルス」 (「小人」) 組織を持っているのに対し、小脳は、より広範囲に分布した投影を備えた、より外積のような構造を保持していますが、内側-外側の地形のある程度の尺度は保持しています ([[@LuMiyachiItoEtAl07]])。
{id=”figure_anatomy” style=”高さ:30em”}
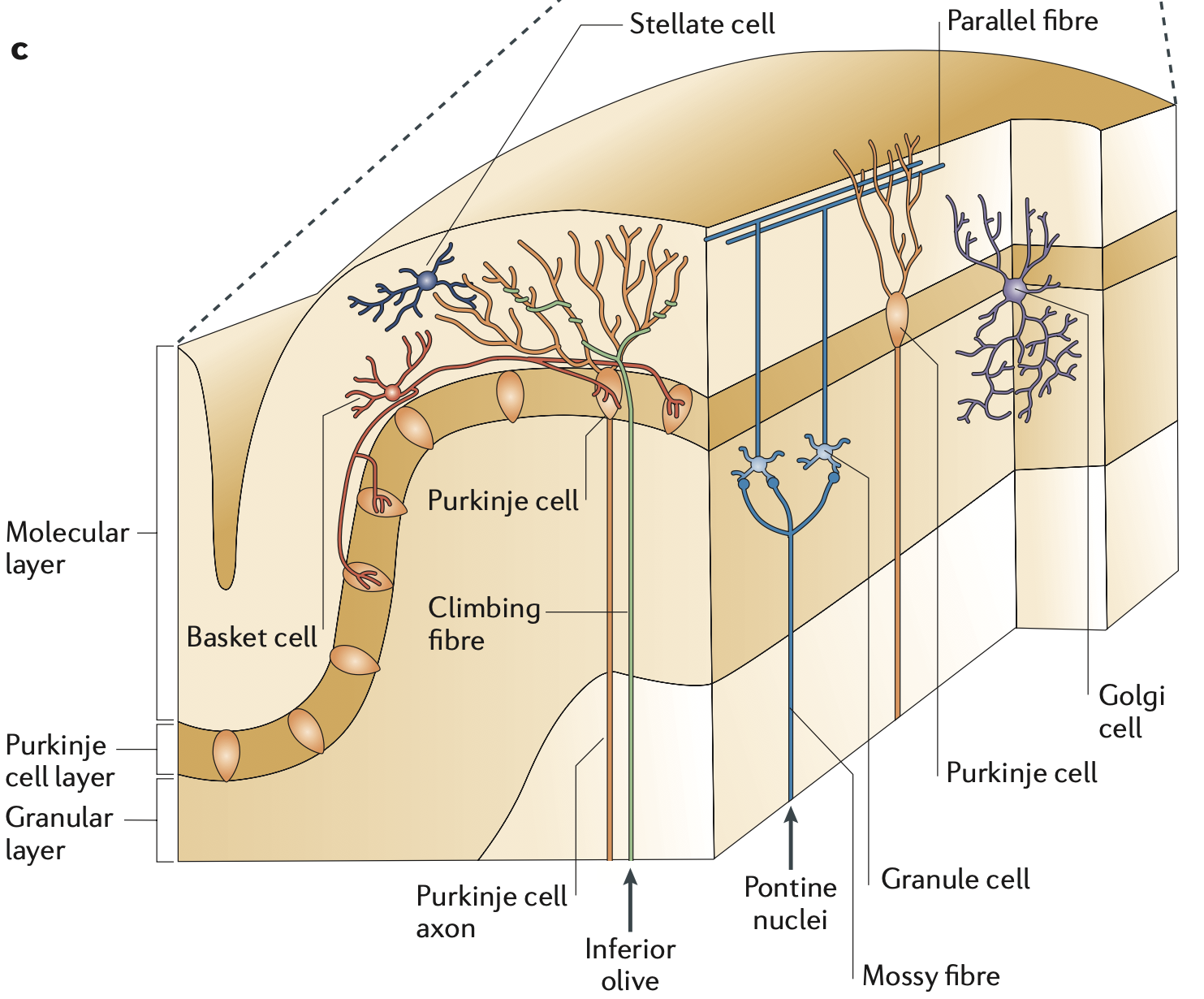
[[@^Ramnani06]] の図 [[#figure_anatomy]] は、小脳の主要な解剖学的特徴を示しており、その多くは他の脳領域と比べて特徴的です。これらの各機能を、以下で開発された機能モデルのコンテキストで詳細に検討します。小脳皮質の基本回路は、さまざまな感覚運動入力(たとえば、橋核から)を数百万の小さな顆粒細胞に伝える苔状線維の入力から始まり、その後、膨大な数の平行線維を皮質からの唯一の出力ニューロンであるプルキンエ細胞に送ります。
プルキンエ細胞は、比較的高い速度 (約 50 Hz) で緊張的に活動し、同様に緊張的に活動する興奮性小脳核出力ニューロン (図示せず) に抑制を送ります。これらは、[[@^Marr69]]、[[@^Albus71]]、および [[@^Ito72]] の元の計算モデルと一致して、プルキンエ細胞の学習を促進することが知られている、正確に 1 つの下オリーブ (IO) ニューロンから _登山線維 _ 入力を受け取ります。
重要なことに、この回路は、小脳全体で複製されるすべてのループにわたる大部分の「並列」処理をサポートしており、異なるループ間の相互作用はほとんどありません。顆粒細胞の平行繊維は、同じ感覚入力信号をこれらのループ全体に効率的に分配しますが、各プルキンエ細胞出力チャネルはほぼ独立しています。これは、他の脳領域、主に [[basal ganglia]]、特に [[neocortex]] が、意思決定と運動動作の階層的組織化をサポートするために、これらのチャネル全体の統合と比較を担当していることを意味します。したがって、小脳は、それ自体は行動を選択したり形成したりするのには適していないが、むしろ他の場所で選択された行動を導くための高次元で調整された感覚基盤を提供するために重要である。
小脳内には、小脳核の興奮性出力ニューロンから皮質の顆粒細胞とゴルジ細胞に至る、興味深い、しかし無視されがちな直接フィードバック投射が 1 つあります ([[@HouckPerson14]]; [[@HouckPerson15]]; [[@AnkriHussonPietrajtisEtAl15]])。これにより、他の小脳経路が学習できるように、小脳の現在の出力を迅速にブロードキャストできます。このフィードバック ループは、これらの小脳核ニューロンにプルキンエ投影を送信する同じモジュールにも入力を提供するため、閉ループ フィードバックと_開ループ_ フィードバックが提供されます。
私たちが採用するモデルの出発点は、小脳が運動動作の感覚効果を予測し、これらの予測された感覚信号を感覚入力の進行中のストリームから「減算」することを学習することによって、運動制御の安定した効果的な基盤を提供するということです ([[@Fujita82a]]; [[@DeanPorrillEkerotEtAl10]]; [[@Cullen23]])。この適応フィルタリング機能は、小脳皮質の唯一の出力を表すプルキンエ細胞から小脳核への大規模な抑制性投射によって直接サポートされています。最終的な効果は、キャンセルされなかった残りの感覚信号が、自分自身の行動の結果を超えた何かを表し、さらなる行動を推進するための適切な基礎となることです。
1 セットの運動動作によって生成される感覚信号のキャンセルをサポートする同じタイプの予測学習メカニズムは、最初の動作の予想される感覚結果に基づいて後続の運動動作を駆動するのにも役立ちます。この典型的な例は、広く研究されている [[#vestibulo-ocular reflex]] (VOR) です。この VOR では、頭の動きによって生成される予測前庭信号が反対方向の矯正眼球運動を駆動し、網膜上で安定した視覚刺激を維持します ([[@ZeeuwHanselBianEtAl98]]; [[@KimpoRinaldiKimEtAl14]])。網膜像を効果的に安定させるには、頭の動きの影響に事後的に反応するのではなく、目の動きが頭の動きの影響を「予測」することが重要です。この関数は、広く議論されている小脳機能の_順モデル_理論 ([[@MiallWolpert96]]; [[@Bastian06]]; [[@TanakaIshikawaLeeEtAl20]]) と一致しています。
多くの理論家が長年にわたって小脳のこれらの機能について議論してきましたが、それらが達成される正確な神経機構は、いくつかの重要な点でやや不明瞭なままでした。さらに、より一般的なもの (例: Marr-Albus-Ito モデルのような基本的なエラー駆動学習、または広く研究されている遅延まばたき条件付けパラダイム; [[@Thompson86]]; [[@RaymondLisbergerMauk96]]; [[@OhyamaMedinaNoresEtAl03]]; [[@NguyenPerson25]]) に基づくフィードフォワード運動制御など、他の多くの機能フレームワークが提案されています。 (例: [[@Kawato99]]、[[@Llinas09]])。
さらに問題を複雑にしているのは、小脳皮質における処理と学習と、小脳核で行われる処理と学習との関係であるが、これは文献では比較的無視されている([[@KebschullCasoniConsalezEtAl24]]; [[@HerzfeldHallTringidesEtAl20]])。小脳核が「統合された」小脳学習の最終的な長期的拠点であり、したがって小脳皮質がより一時的な役割を果たしているという証拠が長年にわたって存在している([[@KassardjianTanChungEtAl05]]; [[@ShutohOhkiKitazawaEtAl06]]; [[@KimpoRinaldiKimEtAl14]]; [[@RaymondMedina18]])。したがって、根底にある神経機構の完全な説明は、小脳核内の回路がどのように独立して完全な予測学習機能をサポートできるのか、そして小脳核内の学習が小脳皮質で起こっている学習とどのように関連しているのかを説明する必要があります。
小脳皮質は確かに、顆粒細胞の数が膨大であるため、さらには小脳核 (CN) のニューロンの数に比べてプルキンエ細胞 (PC) の数の拡大率が大きいため、感覚運動空間をはるかに高解像度で表現します。 [[@^BengtssonEkerotJorntell11]] は、600 ~ 1,200 個の PC が個々の CN 出力ニューロンに集中すると推定しており、マウスの半球あたりの合計 CN 興奮性出力ニューロンの数は約 8,500 個 ([[@Heckroth94]]) であるのに対し、約 200,000 個の PC ([[@Woodruff-Pak06]]) と推定されています。
以下で説明する Nuclear モデルは、最初に小脳核 (CN) の計算に焦点を当てており、CN ニューロン間の接続性と可塑性がどのように適応フィルタリングと順モデル機能の両方を直接サポートできるかを示し、特定の種類の抑制性 CN ニューロンによって駆動される予測学習メカニズムを構築しています。我々は、これらの抑制性CNニューロンが特定の感覚入力チャネルを予測して予期することを学習し、下オリーブ(IO)に投射し、同じ感覚入力から正確に対になった興奮性投射も受け取るのではないかと仮説を立てています。これにより、IO がこれらの対の興奮性入力と抑制性入力の差として感覚予測誤差を直接計算できる明示的な神経メカニズムが提供されます。
ペアの IO 入力は通常、IO 樹状突起上の精巧な glomerulus 構造内に統合されており、_temporally-offset_error 計算 ([[@DeZeeuwSimpsonHoogenraadEtAl98]]; [[@LoyolaHooglandHoedemakerEtAl23]]) を実行するように理想的に構成されているように見える複雑な神経特性を持っています。 IO ニューロンは、この重大なエラー信号を小脳皮質の CN 細胞とプルキンエ細胞の両方に投射し、小脳全体の学習を推進します。
| このエラー信号の特性は奇妙に貧弱であるとして広く議論されていますが ([[@NajafiMedina13]]; [[@ZangDeSchutter19]])、私たちは [[#properties-of-the-io-error-signal | below]] が、より段階的な情報を伝達することを犠牲にして、時間精度を損なうことになるが、正確なタイミングで学習を促進するのに実際には意味があると主張します。 |
したがって、Nuclear モデルの神経機構は、小脳核とさらに大きな小脳皮質内での協調学習の完全な説明を提供します。皮質の学習メカニズムは核の学習メカニズムよりも速く、また崩壊しやすいです ([[@HerzfeldHallTringidesEtAl20]]; [[@KimpoRinaldiKimEtAl14]]; [[@KassardjianTanChungEtAl05]])。対照的に、核の学習はゆっくりと持続します。つまり、海馬と新皮質の相補的学習システムのフレームワーク ([[@McClellandMcNaughtonOReilly95]]; [[@OReillyRudy01]]; [[@NormanOReilly03]]) と同様に、CN は永続的な「ルール」をゆっくりと学習しますが、小脳皮質は「例外」を迅速に学習できます。例外が続く場合、特に初期の発達学習中に、例外は CN に統合されます。
核モデルを提示した後、小脳の特定の寄与がより広範な [[motor]] システムにどのように適合し、柔軟で適応的で有能な運動行動をサポートするかを理解することを目的として、それを文献内の他のモデルと比較します。
核モデル
適応フィルタリング
小脳によって実行される予測キャンセル機能は、適応フィルター ([[@Fujita82a]]; [[@WidrowStearns85]]) として知られています。これは、他の感覚信号に基づいて予期または予測されるものをフィルターで除外する [[predictive learning]] の形式であり、residual 信号は、予期しないものを明確に表す「純粋な」感覚信号を表します。
体の一部、特に頭や目を動かすと、すべての感覚に渡って入ってくる信号に大きな変化が生じます (つまり、感覚の再伝達_)。何らかの方法でこれらの「自己運動」感覚信号を知覚システムから差し引かなければ、世界で賢明に行動することは決してできないでしょう ([[@Cullen23]])。すべてが常に乱流状態にあるため、これらの動くターゲットのすべてに基づいて適切な運動動作を調整することは決してできません。
小脳の進化の歴史は、他の動物における原小脳回路および小脳類似体に基づくこの適応フィルター機能と一致しています ([[@BodznickMontgomeryCarey99]]; [[@MontgomeryPerks19]]; [[@BellHanSawtell08]]; [[@Cisek21]])。哺乳類の小脳の最も古い部分は、基本的に脳幹の一次感覚核 [[vestibular]] 感覚核の不可欠な構成要素であり、体のすべてのレベルで自己生成された運動信号の影響を差し引いて、自己運動生成信号に基づいて予想されるものとの残りの不一致を表す「純粋な」前庭信号を生成することを学習します。
この純粋な前庭信号は、効果的に作用できるものになります。自分自身の行動をすべて考慮した後、さらなる動きの感覚がある場合は、滑ったり、転んだり、何らかの形で押し飛ばされたりしていることがわかります。さらに、この純粋な信号は、残留運動に対抗するためにどのように行動すべきかを正確に示します。視覚、触覚、固有受容などの他の感覚入力と組み合わせると、障害の考えられる原因が何かを把握し、それに応じて運動反応を調整することもできます。
{id=”figure_env” style=”高さ:20em”}
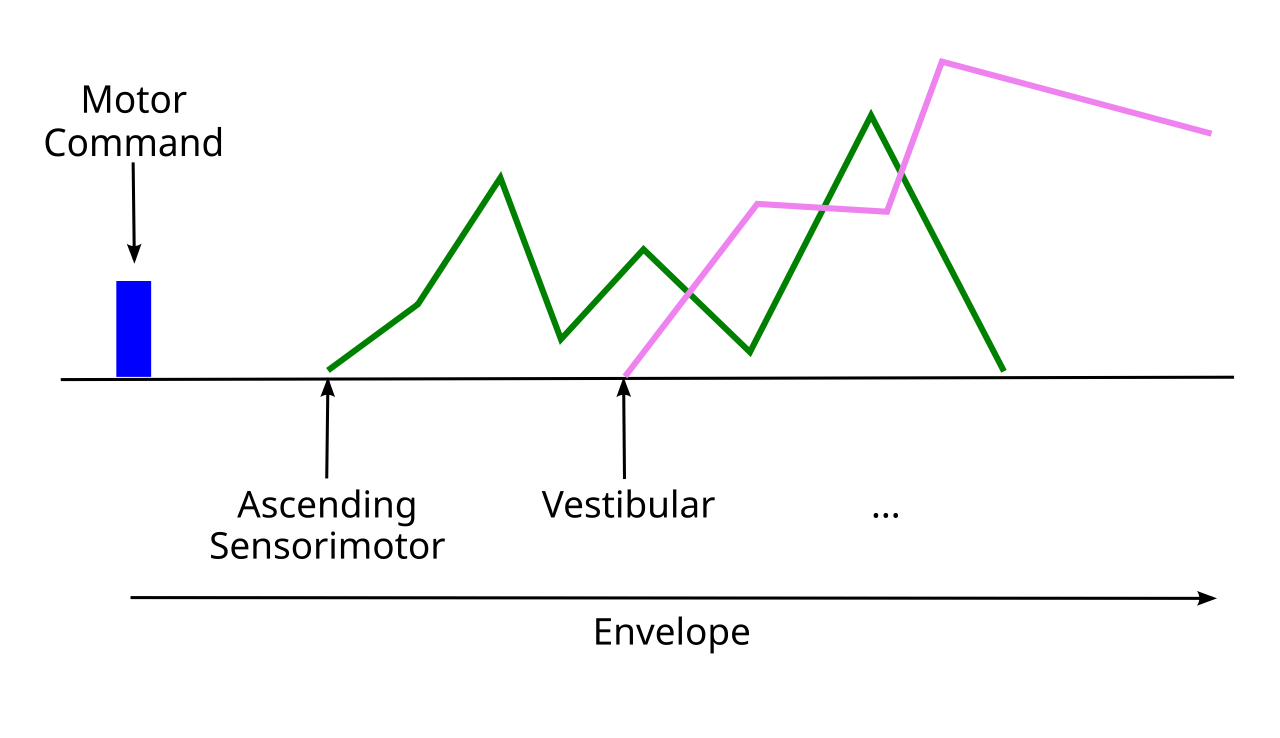
この例から明らかなように、小脳による適応フィルタリングは、賢明な運動動作を生成できるようにするための重要な基盤を提供します。小脳が克服しなければならない主な課題は、運動動作とその感覚伝達の影響に関連するさまざまな時間的遅延を管理することです。頭を動かすと決めたとき、下降運動コマンドの*遠心性コピー**が、これから何が起こるかについての最も早い信号を提供し、すべてを考慮する必要がある後続の信号の複雑なカスケードを開始します ([[#figure_env]])。
脊髄と骨格筋は、効率的な方法で運動指令を実際に実行するためにさらに多くの作業を行うため、下位レベルの脊髄運動経路からバックアップされた実際の上行性運動信号と、筋紡錘線維から伸びている筋肉の現在のレベルに関する関連固有受容信号を取得する必要もあります。これらの信号は、下降するモーター コマンドが比較的弾道的であったとしても、モーターの動作が発生するにつれて時間の経過とともに展開されます。
次に、頭が動き始めると、その動きを反映する前庭系からの信号を受け取り始めます。これらの信号の特定の時空間パターンは正確なモーター パラメーターに依存するため、これはさらに困難な予測問題となります。 [[#figure_env]] に示すように、小脳によって実行される適応フィルタリング ジョブは、最初の下降運動コマンドによって定義される「側頭包絡線」全体を中心に組織化できます。
{id=”figure_cancel” style=”高さ:20em”}
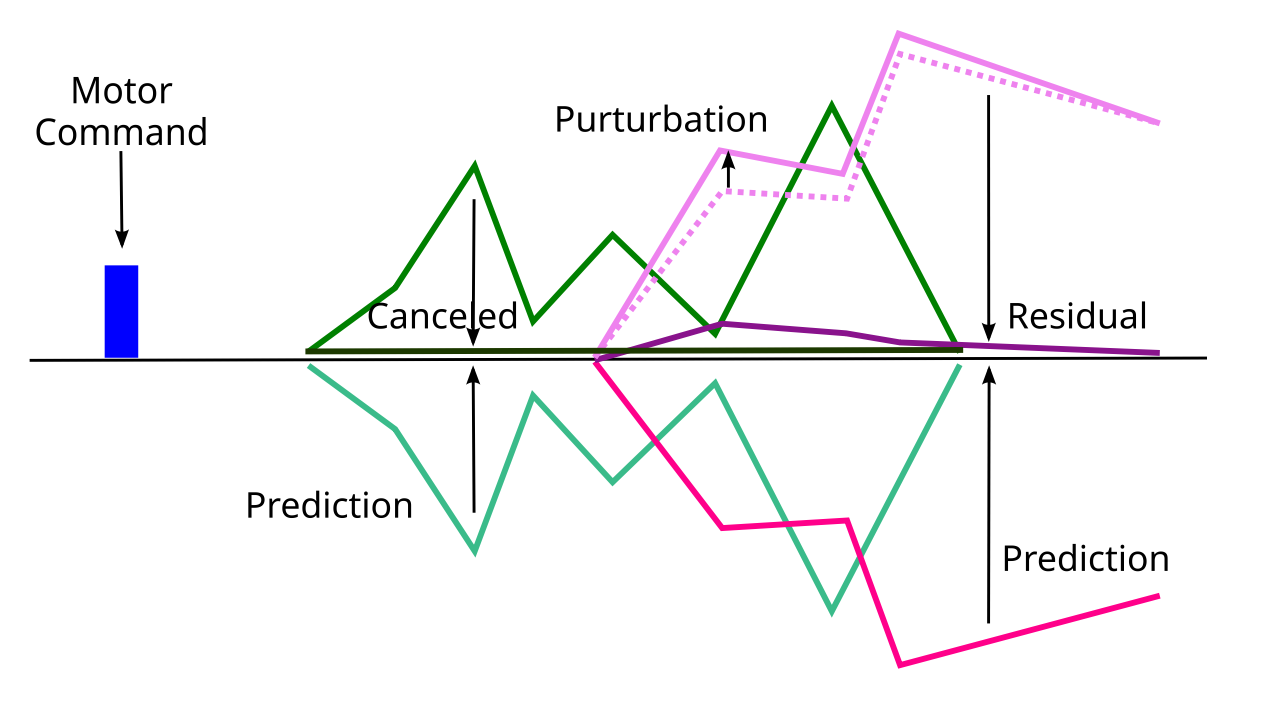
適応フィルターによって実行される中心的な計算は、実際の感覚入力 ([[#figure_cancel]]) から運動動作の感覚結果の学習された予測を減算することです。実際の感覚入力が学習された予測と一致する場合、結果は感覚信号の「キャンセル」になります。ただし、他の予期せぬ出来事 (スリップ、プッシュなど) による感覚信号の摂動がある場合、これは予測によってキャンセルされていない「残留」信号として明確に表示されます。
フォワードモデル
抑制解除プロセスに加えて、学習された感覚予測信号自体は、運動動作の感覚結果に関する予測情報を提供するため、運動制御をサポートする上で直接価値があります。運動制御に使用される感覚結果のこの先取り予測は、順モデル ([[@JordanRumelhart92]]; [[@ShadmehrSmithKrakauer10]]) として知られており、小脳処理の重要な側面として長い間仮説が立てられてきました ([[@MiallWolpert96]]; [[@Bastian06]]; [[@TanakaIshikawaLeeEtAl20]])。
フォワード モデルと適応フィルターの区別は、多くの場合、文献では明確ではありません。これは、どちらも感覚信号の予測学習という同じコア計算を必要とするためです。ここで採用した用語では、順モデルをこれらの予測信号を使用して、予測された状態に基づいて運動動作を直接駆動するものとして定義しますが、適応フィルターはこれらの予測信号を感覚状態から「除去」します。
適応フィルターは、キャンセルを促進するために予測された感覚信号に比例して抑制を強化する必要がありますが、順方向モデル信号は感覚予測に比例するように逆方向に進む必要があります。どちらも、運動動作の遠心性コピーによって引き起こされる予測学習メカニズムによって駆動されるべきであり、すべてをキャンセルすることを常に学習するだけではなく、動作「について」である必要があります。
同じ前庭信号で動作するこれら 2 つの機能の明確な例の 1 つは、前庭信号から減算する頭部の動きの適応フィルタリング ([[@Cullen23]]) と、頭部の動きを補正するために目の動きが生成され、網膜上で視覚刺激を安定に保つ [[#vestibulo-ocular reflex]] (VOR) です ([[@ZeeuwHanselBianEtAl98]]; [[@KimpoRinaldiKimEtAl14]]; を参照)。 [[VOR simulation]])。 VOR では、頭部運動アクションの影響を予測して眼球運動アクションを生成する必要があり、これには順方向モデルの予測が必要です。
感覚信号を予測し、できるだけ早く反応できるようにするために、小脳回路はすべて並行して動作します。したがって、適応フィルタリングとフォワード モデルの機能は、論理的には関連していますが、並列ループを介して動作します。適応フィルタリング回路からの信号をVOR用の別の小脳回路に順次送り返すことによってVORの眼球動作を生成するには遅すぎるであろう。
小脳は、適応フィルタリングと前方モデルの特定の要求を並行してサポートするように組織されています。小脳核は、一般的な予測学習の側面を直接実装し、また、下オリーブ核への入力を介して予測誤差信号を小脳皮質に送信します。重要なことに、この IO エラー信号は、上向き (ゼブリン II 陽性) および **下向き ** (ゼブリン II 陰性; [[@DeZeeuw21]]) として知られる、異なるマイクロゾーンで異なる方向の可塑性を引き起こす可能性があります。これらの異なるマイクロゾーンは、それぞれ適応フィルタリング機能とフォワード モデル機能をサポートできます。
小脳の学習が小脳核の回路に統合されているというかなりの証拠 ([[@KassardjianTanChungEtAl05]]; [[@ShutohOhkiKitazawaEtAl06]]; [[@KimpoRinaldiKimEtAl14]]; [[@RaymondMedina18]]; [[@HerzfeldHallTringidesEtAl20]]; [[@KebschullCasoniConsalezEtAl24]]) を考えると、これらの回路は、適応フィルタリングとフォワードモデル機能。これがどのように機能するかを理解することで、小脳系の多くの機能について明確で斬新な機能的説明が得られます。
原子力回路
{id=”figure_nuclear-forward” style=”高さ:25em”}
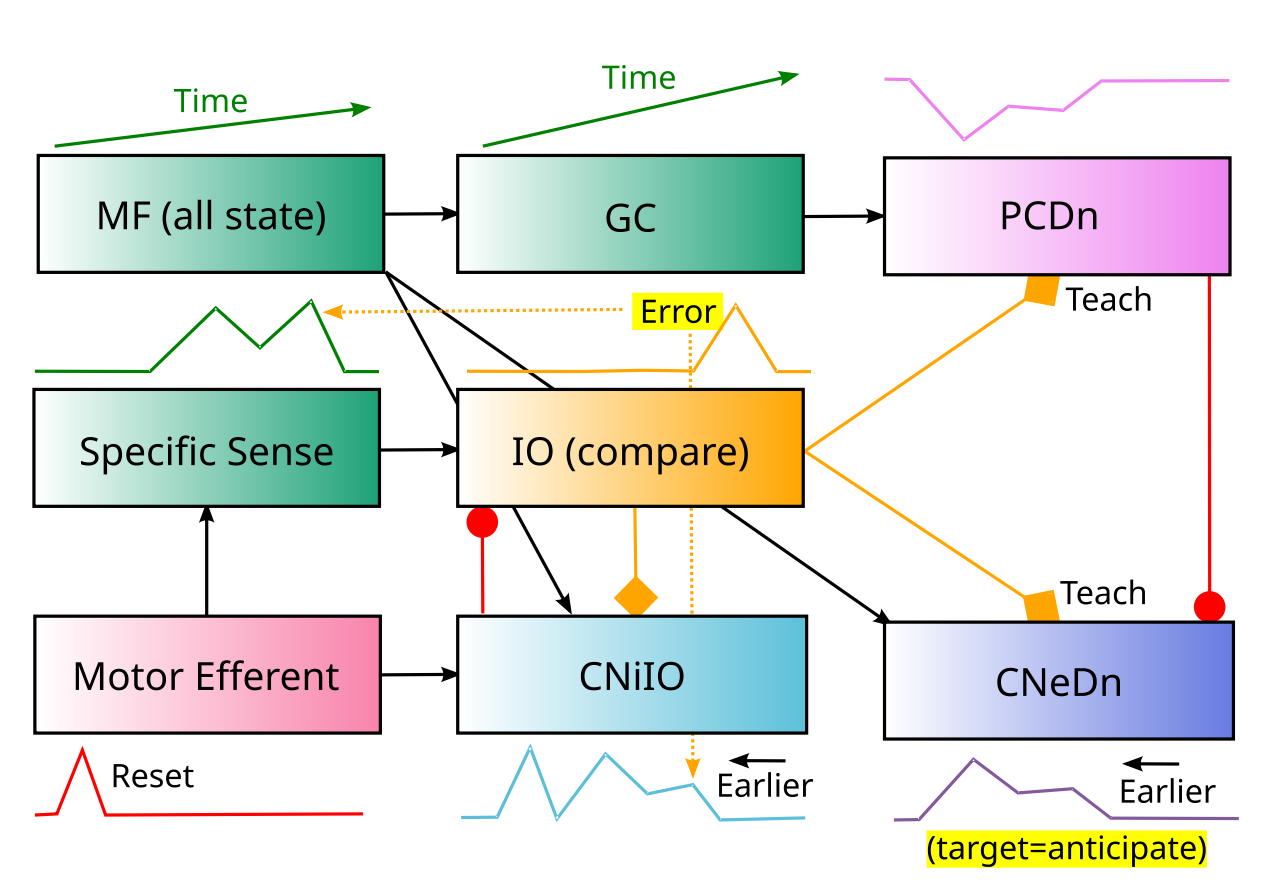
{id=”figure_nuclear-adapt” style=”高さ:25em”}

[[#figure_nuclear-forward]] は、Nuclear モデルに従ってフォワード モデル プロセスを実装できる小脳回路の主要な要素を示し、[[#figure_nuclear-adapt]] は適応フィルタリングのケースを示します。このモデルの各主要要素は文献で文書化され議論されていますが、具体的な詳細はすべて 1 つの一貫したフレームワークに統合されていません。 Nuclear フレームワークの概要を説明した後、[[#Standard models]] との比較ポイントについて説明します。
それぞれの特定の感覚チャネル (例: 前庭系の 1 つの特定の管) には、マイクロゾーン 組織 ([[@Oscarsson79]]; [[@SugiharaShinoda04]]; [[@Oscarsson79]]; [[@SugiharaShinoda04]]; [[@HawkesLeclerc87]]; [[@DeZeeuw21]])。重要な追加の仮説は、特定のマイクロゾーンのIOに投影される同じ特定の感覚入力が、上向きのマイクロゾーンの対応するCNニューロンにも投影され、そこでは運動動作からは予期されない感覚信号を反映する出力活動を駆動するために必要であるということです。
IO 投影は、十分に文書化されているクライミング ファイバーと複雑なスパイク メカニズムを介して、CN ニューロン ([[@vanderWantVoogd87]]、[[@DeZeeuwVanAlphenHawkinsEtAl97]]、[[@RuigrokVoogd00]]、[[@BaumelJacobsonCohen09]]、[[@RowlandJaeger05]]) と PC ニューロンの両方に「今すぐ学習」という教育信号を提供します。この IO 信号は、CNiIO 抑制予測と特定の感覚入力間の不一致を反映しています。 CNiIO 予測は初期の運動遠心性信号によってトリガーされるため、その後の感覚信号よりもおよそ 50 ~ 100 ミリ秒前に発生します。そのため、IO には、初期の予測を後の感覚入力と比較できるようにするための特殊な時間オフセット メカニズムが必要です。
下向きのマイクロゾーンにおける順方向モデル ケースは、完全に初期のタイムスケールで動作し、運動動作の影響を最大限に予測します。対照的に、上りマイクロゾーンの適応フィルタリングの場合は、後の感覚入力信号と同期して抑制のキャンセルを駆動する必要があります。この重要な違いは、アップバウンド経路とダウンバウンド経路の間の遅延の違い ([[@tenBrinkeBoeleSpankeEtAl15]]) と一致しており、CN 内で異なる接続と学習ロジックが必要です。このレベルのマイクロゾーン固有の接続性に関するデータはまだ存在しないため (以下で説明するいくつかの注目すべき例外を除く)、したがって、核モデルはいくつかのテスト可能な予測を生成します。
最初に説明した CN 回路は、適応フィルタリングと順モデル機能の完全な実装を提供します。これは、これが統合された長期的な小脳学習の軌跡であることを示すデータと一致しています ([[@KassardjianTanChungEtAl05]]; [[@ShutohOhkiKitazawaEtAl06]]; [[@HerzfeldHallTringidesEtAl20]]; [[@KimpoRinaldiKimEtAl14]])。無傷の脳のCNニューロンの可塑性に関する証拠([[@MedinaLisberger08]]; [[@KimpoRinaldiKimEtAl14]]; [[@YangLisberger14a]]; [[@HerzfeldHallTringidesEtAl20]])と、スライスにおけるCN可塑性の直接試験([[@AizenmanHuangManisEtAl00]]; [[@PughRaman08]]; [[@PughRaman06]]; [[@ZhangLinden06]]; [[@McElvainBagnallSakatosEtAl10]])。
CNiIO ニューロンはまた、IO 登攀線維複合体スパイクへの影響を介して小脳皮質の学習を促進します。皮質は、アップバウンドおよびダウンバウンドのマイクロゾーンにわたって、より迅速かつ時間的に正確なバージョンのフォワード モデルおよび適応フィルタリング機能を提供できます。 Nuclear モデルの特定のコンポーネントを示した後、モデルで使用される実際の方程式を含め、システムの各要素の学習関数のより詳細な考察が示されます。
特定の感覚および運動入力
Nuclear モデルの重要な要素は、各マイクロゾーンが特定の感覚入力信号 (前庭管など) の処理を中心に編成されており、信号は予測される (順方向モデルの場合) かキャンセルされる (適応フィルターの場合) かのいずれかです。さらに、これらの機能は特定の動作 (例: 頭の回転) に依存しており、この動作は感覚の続発症よりかなり前に発生します。
{id=”table_inputs” title=”CN、小脳皮質への解剖学的入力 (Kebschull et al.、2024) (Med = 内側、PIN = 後部介在、AIN = 前方介在、Lat = 外側)”} |入力 |モダリティ |メッド | PIN |アイン |緯度 |コーテックス | |—————————-|———————|—–|—–|—–|—–|———-| | 脊髄 | | | | | | | |子宮頸部 |首、腕、手 | 写真 首、腕、手++ | ++ | ++ | – | +++ | |胸部 |トランク | ++ | ++ | ++ | – | +++ | |上部腰椎 |ヒップ、脚 | ++ | ++ | ++ | + | +++ | |仙骨、腰椎下部 | 写真足、つま先 | 写真 足、つま先++ | + | +++ | – | +++ | |クラークのコラム |スピンドルファイバー | + | – | + | – | +++ | | 延髄 | | | | | | | |楔形、しなやか |体性感覚 | – | – | – | – | ++ | |三叉神経 | 三叉神経顔の体性感覚 | ++ | ++ | ++ | ++ | +++ | |前庭神経節 | 前庭神経節一次前庭 | – | – | – | – | ++ | |内側前庭 (MVN) |二次前庭 | ++ | – | – | – | ++ | | 網状形成 | | | | | | | |パラメディアン (PPRF) |視線 (SC、前庭) | ++ | ++ | ++ | ++ | +++ | |ギガント、巨大細胞 |姿勢、体幹、調子 | ++ | ++ | ++ | ++ | ++ | |横方向 (LRN) |反射神経 | ++ | ++ | +++ | + | +++ | |赤色核 (巨細胞) |歩行、把握 | 写真– | + | ++ | + | – |
特定の感覚入力信号は、[[#table_inputs]] (すべてではない [[@KebschullCasoniConsalezEtAl24]] から適応) に示されているように、前庭核、三叉神経核、後柱核、延髄、網様体核などのさまざまな脊髄核および中脳核から発生します。これらは一般に IO への入力 ([[@SwensonCastro83]]; [[@BrownChan-PalayPalay77]]) と一致しますが、IO は楔状核と薄核から受信します ([[@BerkleyHand78]]) が、CN は受信しません ([[@DietrichsWalberg87]]; [[@QuyFujitaSakamotoEtAl11]])。
[[#figure_nuclear-adapt]] に示すように、特定の感覚入力 (および後述する MDJ 経路) は、CN の上り適応フィルタリング機能にとって特に重要です。具体的には、感覚入力は CNeUp 出力ニューロンを駆動し、CNiUp および PCUp 阻害によってキャンセルされない予測できない感覚信号を正しく反映する必要があります。
上りエリアが特定の感覚入力を受信していることと一致する証拠がいくつかあります。広いレベルでは、小脳皮質の苔状線維経路を形成する多くの小脳入力もCNに投射しますが、例外もあり([[#table_inputs]])、これは一般にCN投射が高レベルの統合領域ではなく、より低レベルの「基本」感覚経路からであることと一致しています。たとえば、橋核ニューロン (新皮質入力を伝達する) の約 5 分の 1 のみが CN への分岐を持っていました ([[@ShinodaSugiuchiFutamiEtAl92]]) のに対し、下位レベルの側方網様核 (LRN) のニューロンはすべて CN 投射を持っていました ([[@WuSugiharaShinoda99]])。
さらに、CN 投影は一般に小脳皮質への投影よりも焦点が合っています ([[@Sugihara11]]; [[@AndoUedaLuoEtAl20]]; [[@ShinodaSugiuchiFutamiEtAl92]]; [[@WuSugiharaShinoda99]])。さらに、MDJ に強く投影される CN 領域はゼブリン II 陽性、つまり上向きです ([[@Sugihara11]]; [[@WangNovelloGaoEtAl22]] も参照)。
{id=”table_groups” title=”機能群 (杉原、2011)。(v = 腹側、d = 背側、m = 内側、l = 外側、c = 尾側、r = 吻側; FN = 側壁、ICG = 間質細胞群、PIN = 後介在、DN = 歯状、DLP = 背外側隆起、LVN = 側前庭核、DMC = 背内側稜、AIN = 前方介在、MD = 中脳) |グループ |ゼブリン II |入力 | CN |コーテックス |ゾーン |イオ | |———-|———–|———-|—————|—————|———-|———–| |私 | + 上 |中脳 | FNvm | 1+//1+ |あ | c-MAO c | | | | | FN v l | 2+//3+ メートル |アックス | | | | | | ICG v | 2+//3+ l |アックス | c-MAO r | | | | | PIN DL | 4+//5+ | C2 | r-マオ | | | | | DN c | 5+//6+ | D1 | v-PO | | | | | DNrl | 6+//7+ | D2 | d-PO | | IIa | + 上 |前庭 | FNVCM | a+//2+m |あ |ベータc | | | | | FN vc l | a+//2+ l |あ |ベータC-R | | | | | PIN v | 2b+//4+ | X-CX | DMCC、DM c、v-PO cm | | IIb | + 上 |コリキュラー | FN ミッドセンチメートル | α+//2+ |ラテンA | c-MAO m | | | | | FNミッド-C DL | c+//4b+ |ラテンA | c-MAO l c | | | | | FN ミッド C l | d+//5a+ | X-CX | c-MAO r | | Ⅲ | – DN |メリーランド州ソーマ、ベスト | FNrdm | 1-m//1- |あ | c-MAO l c | | | | | FNrdl | 1-l/a-//2- | A1 | c-MAO int | | | | | ICG d | 2a-//3- | X | c-MAO r | | | | | DLPr | 2b-//4a- |ラテンA | c-MAO m cl | | | | | DLP d | c-//4b- |ラテンA | c-MAO cm | | | | | DLP c | d-//5a- |ラテンA | c-MAO int | | | | | PINDM | 3b-//e2- | CX | c-MAO r | | Ⅳ | – DN |体性感覚 | LVN、AICG | 2-//4- | B | d-DAO m c | | | | | LVN、AICG | b+//f+ | B | d-DAO l極 | | | | | DMC (cm AIN) | 3+//エル+ | C1 | v-DAO CL | | | | | AINrm | b-//f- | C1 | v-DAO l | | | | | AIN c | 3-//エル- | C1 | v-DAO c | | | | | AIN l | 4-//5- | C3 | v-DAO rm | | V | + 上 |ビジュアル | v-DN、d-Y |フロック、うなずく | D0 | DC、VLO |
[[@^Sugihara11]] (ラットの位置については [[#figure_cortex]] および [[#figure_cortex-nuclei]] を参照) を改変した [[#table_groups]] は、入力に基づいて異なる上りおよび下りのマイクロゾーンをグループに編成する試みを提供します。このことから、ダウンバウンド (順方向モデル) 領域は入力がより多様で、全体的に体性感覚入力により重点を置いているようです。対照的に、上り方向 (適応フィルタリング) は、前庭および視覚、聴覚の定位信号 (丘陵) に重点を置きますが、体性感覚信号は大幅にフィルタリングされません。
感覚入力のこれらの違いは、運動動作を駆動するために使用される感覚信号の観点から意味があり、感覚信号が他の動作を適切に補償するためにアップバウンド適応フィルタリングが必要です。これらの感覚入力は主に視覚および聴覚による位置特定入力であり、到達または移動したい場所への遠位ガイドを提供します。実際、一連の証拠は、皮質のほとんどの運動野が、視覚に基づいた空間参照フレームを計画に使用していることを示しています(例:[[@BatistaBuneoSnyderEtAl99]])。これらの感覚入力 (目と耳) は頭にあるため、前庭入力によって提供される頭の向きについて知る必要もあります。
対照的に、体性感覚入力は運動動作から生じるフィードバック信号を提供しますが、動作の目標や目標を表すものではありません。したがって、ダウンバウンド フォワード モデル領域でのアクションの調整を予測することは重要ですが、適応フィルタリングでは重要ではありません。
[[#table_groups]] にリストされている中間脳系との接続性は、中中脳接合部 (MDJ) から IO の関連領域への投影に基づいており、これは、CN 出力ニューロンから同じ MDJ 領域への対応する投影と関連付けられていることが知られています ([[@DeZeeuwSimpsonHoogenraadEtAl98]]; [[@WangNovelloGaoEtAl22]]; [[@DeZeeuwRuigrok94]]; [[@DeZeeuwSimpsonHoogenraadEtAl98]]; [[@RutherfordZuk-HarperGwyn89]])。ただし、これは、MDJ 野がこれらの CN 野に特定の感覚入力も送信するという意味ではありません。代わりに、CN 領域は、MDJ ニューロンも受信する他の領域から受信します。これらの領域は一般に非常に広範囲に渡っています ([[@KhalilMansvelderWitter22]]; [[@OnoderaHicks95]])。
たとえば、MDJ 核の 1 つであるダークシュヴィッチュ核は、眼球関連運動皮質野、SNr および GPe ([[basal ganglia]] 出力)、[[superior colliculus]]、および不定帯などの中脳構造を含む、多くの異なる眼球運動関連領域の統合器および調整器として機能します。 (ZI) および_後視蓋前核_ ([[@OnoderaHicks95]])。これらの他の入力は、興奮性または抑制性のいずれかになります。おそらく、IO 信号は、より統合的な MDJ ニューロンによって駆動されます。これは、MDJ ニューロンがより行動に関連したレベルのエラー信号を提供するのに対し、CN 適応フィルタリングは、より要素的な感覚信号に適用するのがより効果的であるためです。
他の MDJ 核には、赤色核、カハール間質核、ベヒテレウ核、フォーレル被蓋野、不定帯、傍筋膜下核、および赤核前網様体 ([[@DeZeeuwSimpsonHoogenraadEtAl98]]) が含まれます。 [[#table_inputs]] では大細胞性赤核 (mRN) が興味深いです。なぜなら、それが他のほとんどの入力とは逆のパターンを示し、介在する CN に投射 (およびそこから受信) するが、小脳皮質には投射しないからです ([[@BeitzelHouckLewisEtAl17]])。脊髄([[@BasileQuartuBertinoEtAl21]])への「赤血球脊髄」モーター投射を駆動する mRN 経路と、MDJ 野の構成要素であり、上で述べたように IO および対応する CN 野と相互接続する小細胞 pRN を区別することが重要です。
赤色核のこれら 2 つの側面の区別は、霊長類では解剖学的に明らかですが、他の種では区別されず、それらがより混合されています ([[@LiangPaxinosWatson12]]; [[@BasileQuartuBertinoEtAl21]])。 pRN の機能は文献では依然として謎に包まれていますが、mRN は四足歩行と把握のための遠位肢の協調的な運動制御を提供することが明確に証明されています。文献における赤核の議論では、これらの異なる経路 (例: [[@WangNovelloGaoEtAl22]]) の機能が混同されている可能性があります。
CNiIO 予測学習
抑制性 (GABA 作動性) CNiIO ニューロンは、興奮性感覚入力によって駆動される学習に基づいて、特定の感覚チャネル活動を予測することを学習します。 IO の「今すぐ学習」入力は、苔状繊維 (MF) 入力で学習をトリガーし、感覚入力を予測するエラー駆動学習をサポートできます。 MF の時変入力のサンプルにより、CNiIO ニューロンは時変する特定の感覚入力の予測を生成できます。
[[synaptic plasticity]] メカニズムは以前のシナプス入力活動の統合を反映しているため、これらの MF から CNiIO への接続は、後続の感覚信号 ([[@RaymondLisberger98]]; [[@WangDenkHausser00]]) を「予測」することを学習します。この予測は、アクションの感覚続発症に先立つ運動遠心性信号による初期活性化と一致しているため、CNiIO の予測活動は予測的なままであり、これはフォワード モデルの機能に必要です。
CNiIO ニューロンは、他の種類の CN ニューロン ([[@KebschullCasoniConsalezEtAl24]]、[[@UusisaariKnopfel11]]、[[@OzcanWangBindaEtAl20]]) よりも安静時の活動が著しく低く、これは、主要な出力 CN ニューロンが行うと考えられているように、特定の感覚入力信号を予測し、感覚系や運動系に対して強直性抑制制御を発揮していないという考えと一致します。
CNiIO ニューロンの学習は、PC 入力の影響を受けると仮説が立てられています ([[@PughRaman08]]; [[@YangLisberger14]]; [[@HerzfeldHallTringidesEtAl20]])。ただし、CNe ニューロンとは異なり、CNiIO ニューロンが PC ニューロンから過度の抑制を受けないことが重要です。これにより、予測信号が減少し、より多くの予測誤差が生じるためです。この要件は、CNiIO ニューロンへの PC 抑制入力は比較的弱く、遠位樹状突起でシナプスを形成する一方、PC から CNe への投射は強く、体細胞近くでシナプスを形成することを示す証拠と一致しています ([[@UusisaariKnopfel08]]; [[@UusisaariKnopfel11]]; [[@DeZeeuwBerrebi95]])。
IO 時間的にオフセットされたエラー信号
CNiIO 抑制性ニューロンは、学習した抑制性投影を、関連するマイクロゾーンの 下オリーブ (IO) ニューロンに送信します。これらの抑制性入力は、多くの場合、複数の軸索と樹状突起が集まる IO 樹状突起上の比較的大きな解剖学的構造である糸球体で、特定の感覚入力からの興奮性入力と収束します。狭い時間枠内で興奮性入力と抑制性入力の時間的にオフセットされた対応を検出することにより、IO 糸球体は_予測誤差_ ([[@Ito13]]; [[@DeZeeuwSimpsonHoogenraadEtAl98]]; [[@DeGruijlBosmanDeZeeuwEtAl13]]) を計算できます。
ラットでは、各 IO ニューロンにはそのような糸球体が約 50 個あり、それぞれが 2 つの興奮性入力と 2 つの抑制性入力を受け取ります ([[@DeZeeuwSimpsonHoogenraadEtAl98]])。各糸球体は、近くにある約 5 ~ 6 個の異なる IO ニューロンを、それらの同期を引き起こすギャップ結合によって互いに接続します。したがって、各特定の感覚入力投影を処理する約 5 ~ 6 個の IO ニューロンがあり、小脳の微小領域全体で同じ特定の感覚チャネルをサンプリングする際に冗長性がある可能性があります。たとえば、約 30 個のプルキンエ細胞の軸索が 1 つの CNe ニューロンに集まり、1 つのプルキンエ細胞は少なくとも 20 ~ 50 個の CN 細胞を神経支配できます ([[@AizenmanHuangManisEtAl00]])。
IO 糸球体のエラー検出ロジックは、固有の樹状突起コンダクタンスをリセットする抑制性入力によって動作します。そのため、興奮性入力は ~50 ~ 100 ミリ秒のウィンドウ内では無効になりますが、そのウィンドウの外側では強く増幅されます ([[@LoyolaHooglandHoedemakerEtAl23]])。したがって、ウィンドウの外にあるいくつかのスパイクでも、IO ニューロンで閾値を超えるバースト活動が引き起こされる可能性があります。 IO 内のこのタイム ウィンドウ オフセットは、CNiIO ニューロンが予測的な予測信号を示すことができるようにするために不可欠です。対応する興奮性入力なしで抑制性入力が発生した場合、ニューロンは対応する時間枠の間抑制されたままになります。このような場合に過剰な予測を修正するための潜在的なメカニズムについては、以下で説明します。
IO ニューロンにおける固有電流の複雑な組み合わせにより、ベースライン活動レベルは約 1 Hz となり、抑制性入力と興奮性入力の相対的なバランスとタイミングがスパイクの確率、タイミング、強度を決定します ([[@TitleyKislinSimmonsEtAl19]]; [[@ZangDeSchutter19]]; [[@GuoUusisaari25]]; [[@YangLisberger14]])。少なくとも、これらの IO スパイクは、他の入力によって決定される学習の定量的な詳細とともに、投影先の CN ニューロンと PC ニューロンの両方の可塑性を活性化する粗い信号を提供できます。
さらに、IO 信号が PC ニューロンの学習に意味のある双方向エラー信号を提供できるという少なくともいくつかの証拠があり、少なくともいくつかの十分に文書化されたケースでは、感覚入力に関連する IO 発火の一貫した有無がシナプス可塑性の反対方向を駆動します ([[@OhmaeMedina15]]; [[@YangLisberger14a]])。
また、ギャップ接合による複数の IO ニューロンの同期がエラー信号の有効性の重要な要素である可能性が高く、集団全体での同期した活性または抑制により、より段階的な双方向エラー信号が提供されます ([[@HoangTsutsumiMatsuzakiEtAl23]]; [[@GiessenKoekkoekDorpEtAl08]])。このギャップ結合の接続性は、時間エンベロープ ([[#figure_env]]) を定義する共通の初期運動信号の周りに関連するニューロンのグループを組織するメカニズムを提供する可能性があります。
CNiIO 投影が学習の進行に伴う PC ニューロンの IO 発火と複雑なスパイクの減少に関与していることを示す直接的な証拠があります ([[@KimKrupaThompson98]]; [[@Ito01]])。さらに、この経路は条件付き学習におけるブロック効果の原因であることが示されており、これはデルタ ルールのようなメカニズムにおける予測誤差信号の除去の兆候です ([[@Fanselow98]]; [[@KimKrupaThompson98]])。
PC ニューロンに対する IO スパイクの影響 ([[@Ito01]]、[[@CoesmansWeberDeZeeuwEtAl04]]、[[@GaovanBeugenDeZeeuw12]]、[[@SilvaRamirez-BuriticaPritchettEtAl24]]) については十分に文書化されているのに比べ、IO 入力が CN ニューロンでの学習をどのように促進できるかについてはほとんど知られていません。スライスにおける CN 可塑性の関連研究 ([[@AizenmanHuangManisEtAl00]]; [[@PughRaman08]]; [[@PughRaman06]]; [[@ZhangLinden06]]; [[@McElvainBagnallSakatosEtAl10]]) は、覚醒して行動する動物にはしっかりと存在しない抑制後のリバウンドバースト (つまり、[[in activo]]; [[in activo]]; [[in activo]]; [[@HoebeekWitterRuigrokEtAl10]]; [[@BengtssonEkerotJorntell11]])。たとえば、[[@^BengtssonEkerotJorntell11]] は、CN ニューロンに対する自発的な IO バックグラウンド スパイクの影響を記録し、「巨大な」IPSP を生成しましたが、リバウンド発火の兆候は見つかりませんでした。
さらに、既存の研究では、核モデルによる可塑性の重要な部位である、より小さく、より静止した CNi ニューロンではなく、より大きく、緊張的に活動する CNe ニューロンをサンプリングする可能性があります。これらの CNi ニューロンは遠位の弱い PC 入力のみを受け取るため、それほど強く抑制されず、したがって強いリバウンド活動を示す可能性はさらに低くなります。
データが限られているにもかかわらず、IO 活動が CN ニューロンに重大な影響を与えるという一連の証拠 ([[@HoebeekWitterRuigrokEtAl10]]、[[@BlenkinsopLang11]]、[[@BaumelJacobsonCohen09]]、[[@RowlandJaeger05]]) と、覚醒している動物の CN ニューロンに対する IO 入力による短期可塑性効果の証拠に関する少なくとも 1 件の報告 ([[@GruartBlazquezPastorEtAl94]]) があります。
CNeDn 前方モデルの先行発射
forward model 機能はダウンバウンド マイクロゾーン ([[#figure_nuclear-forward]]) で発生し、IO ニューロンの感覚予測エラーが CNeDn CN 興奮性出力ニューロンでの学習を促進します。 CNeDn ニューロンは、後で到着する特定の感覚入力を正確にキャンセルする必要がないため、できるだけ早く発火して、以前の動作の予測された効果を予測してその後の運動動作を形成するための時間を増やすことができます。これは、ダウンバウンド順モデル経路の主な仮説上の計算上の利点です。
IO 登攀線維は CN に投射し、CNeDn ニューロンへの MF 入力からの学習を促進し、必要な興奮性活動を生成します。これは、前述したように、シナプス統合プロセスと学習の相互作用によって予測できます ([[@RaymondLisberger98]]; [[@WangDenkHausser00]])。これはさまざまな理論 ([[@PughRaman08]]; [[@YangLisberger14]]; [[@HerzfeldHallTringidesEtAl20]]) と一致しています。
興味深いことに、CNiIO ニューロンから橋核および中脳/脳幹運動野への直接投射もいくつかあり ([[@JuddLewisPerson21]])、それによって抑制性の予期信号を送り、興奮性の CNeDn 信号を補完する可能性があります。
CNeUp 適応フィルタリング
小脳の初期の進化的機能は適応フィルタリングであると考えられていますが、運動動作とそれに続く感覚入力の間の時間的オフセットに対処するために、いくつかの追加のメカニズムが必要です(より単純な脳や小型動物では、このオフセットはおそらくはるかに短かったでしょう)。 [[#figure_nuclear-adapt]] に示すように、CNiUp 抑制性介在ニューロンは、CNeUp 興奮性出力ニューロンが予期しない感覚信号のみを伝達するように、入力される感覚信号と正確に一致する後の感覚予測を学習する必要があります。
したがって、核モデルの CNiUp 局所抑制性ニューロンは苔状線維入力を受け取り、IO 教示信号を使用してこれらのシナプスを適応させ、CNeUp 出力ニューロンへの直接的な抑制性投影を介して特定の感覚入力を正確にキャンセルします。
{id=”figure_adapt-mdj” style=”高さ:25em”}

興味深いことに、上向きの CNe ニューロンから MDJ (中脳結合部) ニューロンへの興奮性投射は、対応する CN マイクロゾーン ([[#figure_adapt-mdj]]; [[@DeZeeuwSimpsonHoogenraadEtAl98]]; [[@WangNovelloGaoEtAl22]]) に投射する IO ニューロンに投射され、最終的な CNeUp 信号に基づく直接エラー フィードバック学習のメカニズムを提供します。この信号に残留する摂動は IO スパイクを引き起こす可能性があり、これにより CNiUp ニューロンのさらなる学習が促進され、興奮性感覚入力がより適切にキャンセルされるはずです。これは、これらの一見謎に満ちた MDJ 領域に重要な機能を提供します。
プルキンエ細胞
IO 予測誤差信号は プルキンエ細胞 (PC) の学習を促進し、GABA 阻害性単純スパイクの一定の流れを CNe 出力ニューロンに送信します。したがって、核の観点から見ると、これらの PC ニューロンは CNi ニューロンと同じ機能を持ち、PC ニューロンにおける上り対下りの学習ダイナミクスは、上記の CN 内学習メカニズムと一致します。
{id=”図アップダウン” スタイル=”高さ:45em”}
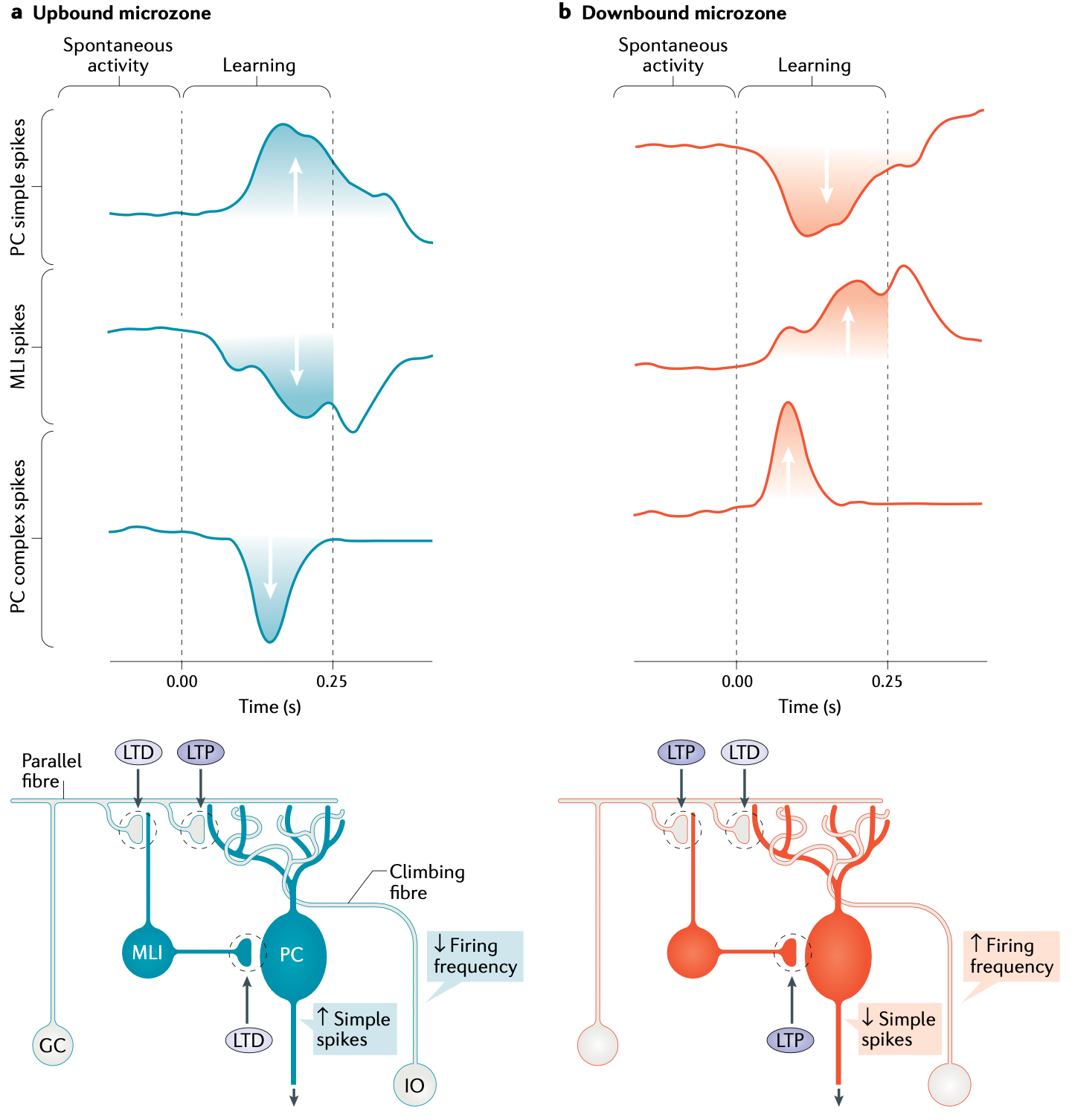
各 IO ニューロンは、登攀線維 (CF) 軸索出力を送信し、(CN 経路に加えて) PC に到達します。各 IO ニューロンは CF 出力を約 10 個の PC ニューロンに送信しますが、各 PC ニューロンは 1 つの IO ニューロンからのみ受信し、PC ニューロンあたり約 300 のシナプスがあります ([[@Ito84]])。 IO スパイクは、PC ニューロンで強力な_複雑なスパイク_ アクティビティ パターンを駆動し、これらのニューロン ([[@ItoKano82]]、[[@CoesmansWeberDeZeeuwEtAl04]]、[[@GaovanBeugenDeZeeuw12]]、[[@SilvaRamirez-BuriticaPritchettEtAl24]]) での学習をトリガーし、学習の方向は [[#figure_up-down]] に示すようにマイクロゾーンによって決定されます。
適応フィルタリングのコンテキスト (つまり、upbound マイクロゾーンの場合) では、予測不可能な感覚興奮を反映する IO 駆動の複雑なスパイクが PCUp ニューロンで LTP (長期増強) を駆動し、CNeUp ニューロンに送信される抑制量が増加し、それによって感覚入力からの追加の興奮性駆動のバランスが取れ、感覚信号がキャンセルされます。
下向きのマイクロゾーンにおける順方向モデルのケースでは、IO 予測誤差信号が PCDn ニューロンのシナプス可塑性の反対の符号を駆動し、その結果、CNeDn ニューロンに正味の脱抑制効果が生じ、それが予測的な運動行動を引き起こす可能性があります。 PC 可塑性のこの反対の符号は、IO 駆動の複雑なスパイクがあるときに発生する LTD に対応します。これは、[[@^Albus71]] および [[@^Ito72]] 運動学習理論によって予測される可塑性のパターンです。
この形態の PC LTD は広範囲に実証され、分析されています ([[@Ito01]]; [[@CoesmansWeberDeZeeuwEtAl04]]; [[@GaovanBeugenDeZeeuw12]]; [[@YangLisberger14a]]; [[@SilvaRamirez-BuriticaPritchettEtAl24]])。順方向モデル学習に必要なこの下向き方向が、可塑性の最も広く研究されている方向であることは注目に値します。これはおそらく、下向き方向がサポートする運動動作が、上向き学習によってサポートされる適応フィルタリングよりも研究がはるかに簡単であるためです。
小脳皮質が可塑性の各方向 ([[@DeZeeuw21]]) に専用のマイクロゾーンを持っているという事実は、適応フィルタリングとフォワード モデルの機能に対する差動接続の要求と一致しています。さらに、各マイクロゾーンで一方向の可塑性が優勢であるという事実は、IO 登坂線維トレーニング信号のやや特殊な性質を反映しています。この信号は本来非対称であり、感覚入力が CNiIO 阻害予測を超えた場合にのみ複雑なスパイクを発火します。したがって、このシステムは、肯定的な予測エラー (予期しない感覚結果の発生) に迅速に対応するように構成されていると思われます。
したがって、期待された予測が起こらなくなった場合、または感覚的な予測強度が実際のレベルを超えた場合に何が起こるかについては疑問が残ります。経験的に、PC と CN の可塑性は双方向であるという証拠 ([[@BrooksCarriotCullen15]]; [[@BrooksCullen13]]) が、一般に「負の」方向 (つまり、下りの場合は LTP、上りの場合は LTD) では遅くなるという複数のレベルの証拠があります。
実際、相補的な方向を、小脳核内で起こるより遅い獲得プロセスに適応するためにも必要な遅い減衰メカニズムと区別することは困難です。これは、小脳皮質による急速な学習から小脳核での遅い統合までのシステムの統合と一致しています。
苔むした繊維の入力
| 苔状線維 (MF) 入力は、小脳の予測発火を駆動するための「ソース」情報を提供します。したがって、行動のその後の感覚的および運動的結果を「確実に予測」する情報を提供する必要があります。最も明白なソースは、そもそも動作を駆動する実際のモーター コマンドです。これらの下行運動指令は主に、運動前頭葉 [[neocortex | cortex]]、[[superior colliculus]] (SC)、および [[basal ganglia]] (BG) の 3 つの主要な脳構造から発生し、それぞれが多くの苔状線維の供給源となる橋核 (PN) に広範囲に投射されます。運動皮質の投射は主に PN を介しますが、SC と BG はそれぞれ MF 入力を駆動するための複数のルートを持っています。 |
SCは、PN(QQPH00000PHQQ)に直接投影し、また、例えばMDJの文脈で上述したように、同様にCNに投影する他のほとんどの中脳運動構造にも投影する。進化的に形成された感覚運動行動の主要なソースとして、SC 入力は、運動コマンドの運転行動に関する重要な初期予測信号を提供します。
大脳基底核からの投射は、視床下核 (STN) ([[@BostanDumStrick10]]) および視床筋膜傍核 (PF) ([[@MariniPiancaTredici99]]) から始まり、BG 出力の主な表現となります。これら 2 つの領域は、BG 活動のポジティブな表現を伝えることができる主要な興奮領域を表しており、STN は BG 処理サイクルの開始時に非常に初期の活動を示します (新皮質および SC 入力によって駆動されます; [[@CoizetGrahamMossEtAl09]])。対照的に、PF は BG の出力を反映し、SNr と GPi からの抑制性投射を介して、SC および中脳運動野を含む広範囲の皮質下の標的に直接伝達されます。
顆粒細胞
小脳皮質には、非常に多数の小さな 顆粒細胞 (GC) も含まれており、これらの細胞は MF 入力を受け取り、独特の平行線維軸索を介して PC に興奮を送り、各 PC は 150,000 以上の平行線維入力を受け取ります。これらの GC が多数あるため、小脳が人間の脳のニューロンの総数の約 70% を占めることになります。これらの GC は、一部の領域の抑制性ゴルジ細胞やその他の特殊な細胞型とともに、PC ニューロンの新しい時間的発火パターンを迅速に学習するのに特に有用な、非常に高次元の時間変化表現を提供すると考えられています ([[@Marr69]]; [[@BuonomanoMauk94]]; [[@MaukBuonomano04]])。
#### まとめ
要約すると、Nuclear モデルは、小脳回路の精巧で特殊な性質が適応フィルターとフォワード モデルの機能を実行するのにどのように理想的であるかを示しており、これは長年の主要な仮説 ([[@Fujita82a]]; [[@MiallWolpert96]]; [[@Bastian06]]; [[@DeanPorrillEkerotEtAl10]]; [[@TanakaIshikawaLeeEtAl20]]; [[@Cullen23]])。
感覚信号をキャンセルするための 2 つの異なる経路、つまり CN 内の直接経路と小脳皮質を通る間接経路が存在することにより、なぜ 2 つの別個の経路が有用なのか、そしてそれらはどのように相補的な機能を発揮するのかという疑問が生じます。 [[hippocampus]] ([[@Marr71]]) と小脳 ([[@Marr69]]) の両方に関する David Marr の独創的なアイデアからインスピレーションを得た 1 つの仮説は、小脳皮質の非常に高次元の表現空間を (GC を介して) 使用することで、システムが事前の学習への広範な干渉を受けることなく、新しい時空間発火パターンを迅速に学習できるようにするというものです。
さらに、CN システムは最も信頼性が高く安定した形式の感覚予測をゆっくりと学習することができますが、小脳皮質はより状況に依存した特異な予測を急速に学習することができ、時間の経過とともに減衰する可能性があります ([[@HerzfeldHallTringidesEtAl20]]; [[@KimpoRinaldiKimEtAl14]]; [[@KassardjianTanChungEtAl05]])。つまり、海馬と新皮質の相補的学習システムのフレームワーク ([[@McClellandMcNaughtonOReilly95]]; [[@OReillyRudy01]]; [[@NormanOReilly03]]) と同様に、CN は「規則」をゆっくりと学習しますが、小脳皮質は「例外」を迅速に学習できます。
CN 経路は皮質経路 ([[@GarciaSteeleMauk99]]) に比べてタイミングが正確ではないという証拠があり、これは小脳皮質で形成される高次元表現と一致しています。各 PC は約 150,000 の顆粒細胞入力からの入力を統合するため、より限定された CNiIO ニューロンの樹状突起よりも多くの多様な刺激入力にわたって新規の任意の関連付けを形成できます。したがって、中枢神経系はより粗いタイミングを学習する一方、小脳皮質は細かいタイミングを学習すると考えられます。このきめの細かいタイミングが持続的に関連する場合、継続的なエラー信号が皮質経路への寄与を維持し、時間の経過による安定化プロセスによりこれらの接続が長期にわたって永続的になる可能性があります。
多くの研究は、小脳皮質における迅速な 1 試行学習と一致しています ([[@MedinaLisberger08]]; [[@KimpoRinaldiKimEtAl14]]; [[@YangLisberger14a]]; [[@HerzfeldHallTringidesEtAl20]])。これらの研究は、単一の試験で発生する最も強力なシナプスの変化が急速に減衰することも示していますが、小脳皮質シナプスにおける長期にわたる変化のかなりの証拠とも一致しています([[@SilvaRamirez-BuriticaPritchettEtAl24]]; [[@Ito01]]; [[@WangNakadateMasugi-TokitaEtAl14]])。
スライスで実施されるほとんどのシナプス可塑性研究で使用される刺激パラダイム ([[@Ito01]]; [[@Ito13]]) と、覚醒行動動物における急速だが一時的な変化を示す刺激パラダイム ([[@MedinaLisberger08]]; [[@KimpoRinaldiKimEtAl14]]) の間には、重大な不一致があり、これが広範な小脳におけるこれらの動態に焦点が当てられていないことを説明している可能性があります。シナプス可塑性の文献。
学習ルール
小脳系のほぼすべてのシナプスに [[synaptic plasticity]] の生物学的証拠があります ([[@HanselLindenDAngelo01]]; [[@Carey11]]; [[@GaovanBeugenDeZeeuw12]])。 [[#figure_nuclear-forward]] および [[#figure_nuclear-adapt]] のロジックに基づくこの可塑性にとって最も重要なシナプスは、CNiIO、CNiUp、および PC ニューロンへの MF 苔状線維入力であり、それぞれが位相的で予測的な活動パターンを学習する必要があります。
この分野では、CN での遅い学習を「システム統合」するには、PC ニューロンでの初期のより速い学習が必要であるという一般的なコンセンサスがあります ([[@RaymondMedina18]]; [[@HerzfeldHallTringidesEtAl20]])。しかし、この結論を裏付ける関連研究のそれぞれには、この点に疑問を残す問題があり、CNでの学習はPC入力から独立して進行できるが、これらのPC入力が存在する場合には学習を促進する可能性があることを示唆しています。
おそらく最も強力に見えるデータは、[[@^GarciaSteeleMauk99]] パラダイムで学習後に大脳皮質に損傷を与え、この損傷がその後の新しい CS の獲得を妨げることを発見した [[@^GarciaSteeleMauk99]] からのものです。ただし、[[@^KimKrupaThompson98]] で判明したように、これは実際には以前の CS-US コンディショニングによるブロック効果によるものである可能性があります。
[[@^ZeeuwHanselBianEtAl98]] による別の研究では、遺伝子ノックアウトを使用して、[[synaptic plasticity]] に関与する重要なプロテインキナーゼを PC ニューロンで選択的に無効にしました。彼らは、これが PC と CN の両方で [[#vestibulo-ocular reflex]] (VOR) の適応をブロックすることを発見しましたが、別の研究で大脳皮質 ([[@KassardjianTanChungEtAl05]]) での学習にのみ影響を与えることが示された短時間のトレーニング (1 時間) のみを使用しました。
速い解離と遅い解離と一致して、[[@^KassardjianTanChungEtAl05]] は 3 日間のトレーニング後にのみ CN で皮質独立学習を発見しました。さらに、[[@^ZeeuwHanselBianEtAl98]] は、ノックアウト マウスでは標準的な VOR とかなりの運動行動が損なわれていないことに注目し、PC の可塑性が CN の発達期間にわたる遅い学習にとって重要ではないという強力な証拠を提供しました。
[[@^ShutohOhkiKitazawaEtAl06]] による別の研究では、さまざまな操作が使用され、IO の不活化により長期的な VOR 適応が著しく損なわれることがわかりました。これは、IO が CN ニューロンの学習に重要であるという考えと一致しています。小脳皮質の不活性化と薬理学的操作は一貫してIOよりも効果は低いが、それでも重大な効果があり、これはCN内での学習におけるPCからCNへの投影の追加の役割と一致している。これは [[@^ZeeuwHanselBianEtAl98]] 研究と矛盾する可能性があり、これらの不一致を解決するにはさらなる研究が必要です ([[@Ito13]] も参照)。それにもかかわらず、CN ニューロンは IO 入力と連動して独立して学習でき、PC 入力は自身の高速学習に基づいて追加の学習信号を提供すると仮定するのが合理的と思われます。
予測デルタ ルール
計算レベルでは、単純な [[error-backpropagation#delta-rule]] 関数は、受信ニューロン y (CNiIO ニューロンの場合など) の活動における感覚入力信号 s を予測する方法を学習するための数学的観点から最適です。
{id=”eq_delta-w” title=”デルタ ルール”} \(\デルタ w = (s - y) x\)
ここで、x は (苔状ファイバー入力からの) 送信アクティビティです。
この方程式は、感覚値と予測の間の不一致に基づいた双方向の学習を必要とします。予測が低すぎる場合は、シナプスの重みを増加 (LTP) する必要があり、逆の場合はシナプスの重みを減少 (LTD) する必要があります。一般に、CN ニューロンにおける双方向のシナプス可塑性については直接的な証拠があります ([[@AizenmanHuangManisEtAl00]]; [[@PughRaman08]]; [[@PughRaman06]]; [[@ZhangLinden06]]; [[@McElvainBagnallSakatosEtAl10]])。
ただし、小脳の「予測」という性質により、さらなる課題が生じます。これは、学習ルールに時間を追加すると明らかになります。$\epsilon$ は、予測 $y$ が予測する事前のミリ秒数 (例: 80 ミリ秒) を表します。
{id=”eq_delta-w-t” title=”時間ベースのデルタ ルール”} \(\デルタ w_t = (s_t - y_{t-\epsilon}) x_{t-\epsilon}\)
(同様に、$\epsilon$ を [[#eq_delta-w-t]] 内のすべての時間インデックスに追加できます)。
したがって、誤差項 ($s_t - y_{t-\epsilon}$) は、予測時間オフセットに敏感である必要があるため、時間 $t$ での予測活動は、たとえば 80 ミリ秒後に到着する (つまり、後の入力を予測する) 感覚入力とほぼ同じになります。入力における自然なトレースのような積分プロセスは、送信入力 ([[@RaymondLisberger98]]; [[@WangDenkHausser00]]) の $x_{t-\epsilon}$ 係数を処理できますが、時間にわたってオフセットされる誤差項を計算するには、実際にはある種の遅延コンパレータ プロセスが必要です。
上で説明したように、IO ニューロンは、まさにこの種の時間的にオフセットされた比較機能を実行できる特殊な神経機構を備えています。この機能では、約 50 ~ 100 ミリ秒後に到着する興奮性感覚入力の影響をブロックするには、CNiIO ニューロンからのより早い抑制性予測入力が必要です。これは、CNiIO および他の CN ニューロンでの学習は、PC ニューロンと同様に、IO ニューロンからのエラー信号に依存する必要があることを示唆しています。これは、IO から CN への側副投影の存在と一致します ([[@vanderWantVoogd87]]; [[@DeZeeuwVanAlphenHawkinsEtAl97]])。したがって、この IO 信号の計算特性を理解する必要があります。
IO エラー信号のプロパティ
IO が小脳の学習を促進するエラー信号を提供するという広く受け入れられている計算レベルの考えは、IO ニューロンにおける神経発火の実際の特性と一致させるのが困難でした ([[@NajafiMedina13]]; [[@ZangDeSchutter19]])。それらは、約 1 Hz の低いレートで定期的に起動しますが、これは一見非現実的なベースライン レベルです。このレベルは、起動レートの低下を意味のあるダイナミック レンジの多くを提供するには低すぎますが、エラー ([[@LlinasLangWelsh97]]) を表す IO アクティビティの S/N 比を低下させるのに十分な高さです。 IO アクティビティがエラーの発生を意味する場合、毎秒誤ったアラームが発生します。しかし、この 1 秒に 1 回の IO 起動がない場合、この不在を意味のある信号として解釈するには、役に立たないほど広いタイミング ウィンドウが提供されるだけです。
トニック バックグラウンド スパイクの問題を軽減する可能性があるものとして、実際の IO エラー信号に関連する発火パターンがベースラインの 1 Hz の発火とは異なるという証拠があります。ある一連の研究では、IO ニューロンの振動位相と単一の初期 IO 活動電位 ([[@MathyHoDavieEtAl09]]) によって引き起こされる小さなバーストの数との関係が示されており、これにより信号をバックグラウンドから識別できる可能性があります。最近では、刺激入力が互いに約 100 ミリ秒以内に 1 対の IO スパイクを駆動することが示されており、その結果、通常はバックグラウンド レート ([[@TitleyKislinSimmonsEtAl19]]) によって生成される単独のスパイクと比較して、PC ニューロンのシナプス可塑性がより強固になります。
ただし、これらのペアの IO スパイクは、最近の in vivo カルシウム イメージング研究では観察されず、代わりに、1 Hz のベースライン ([[@GuoUusisaari25]]) と比較して、刺激駆動型の IO 活動について複数の IO ニューロンにわたるより同期的な集団活動が示されました。いずれにしても、PC 活動が双方向に変調されていることを示す強力な生体内神経記録証拠 ([[@CoesmansWeberDeZeeuwEtAl04]]; [[@GaovanBeugenDeZeeuw12]]; [[@KimpoRinaldiKimEtAl14]]; [[@YangLisberger17]]) を考えると、真のエラー信号がバックグラウンド スパイクとは異なる影響を与える何らかの方法がある可能性が高いと思われます。
IO エラー信号がなぜそのような特性を持つのかを理解するには、2 つの重要な考慮事項が役立ちます。まず、学習信号の正確な「タイミング」が最も重要であり、タイミングの精度は、時間の経過とともに展開される生化学プロセスの段階性と直接矛盾します。スパイクが 1 つだけある場合は、そのスパイクが到着した時点で学習プロセスを即座に開始し、その時点の状態を反映できます。ただし、段階的な量を通信するには、複数のスパイクを時間をかけて送信する必要があります。つまり、段階的な量が確立されるまでに、潜在的に変動する時間が経過し、システムの残りの部分の状態が移行することになります。
この 1 つの正確なタイミングのスパイクの場合は、誤差信号が非対称であることも必要です。この 1 つのスパイクには 1 ビットの情報しかなく、ターゲット値が十分に予測されていない場合、または予測がターゲット値を超えた場合にそれを知らせることができます。これら 2 つのうち、予期せぬ感覚入力の開始を知らせることの方が明らかに重要です。
第 2 に、CN ニューロンと PC ニューロンの両方での学習には、それぞれの場合に異なる理由により、実際には双方向または段階的なエラー信号は必要ありません。 CN ニューロンの場合、学習が非常に遅いため、各学習試行での単一の小さな増分で十分であり、各試行での段階的信号の追加値は最小限です。 PC ニューロンの場合、学習は迅速であり、重要なことに、IO の起動を制御する CNiIO 予測を実際に変更することはありません。したがって、誤差は自動的に飽和する必要があり、誤差の段階的な値は実際には意味がありません。
どちらのニューロン タイプでも、双方向エラー信号の代わりに減衰 (CN ではゆっくり、PC ではより速く) を使用できるため、双方向信号は必要ありません。 CNiIo 予測ニューロンの減衰により、下限値がテストされるとシナプス強度が再び上昇するエラー スパイクが時折発生しますが、PC ニューロンの減衰により、システムはより迅速に遅い CN 学習にフォールバックできるようになります。これはいずれの場合も重要です。バックグラウンドの 1 Hz のスパイクが、ゆっくりとした減衰プロセスを駆動する機会を提供している可能性があります。
したがって、これらの考慮事項を考慮すると、IO エラー信号がより意味のあるものになり始めます。さらに、1 Hz のバックグラウンド発射レートでさえ、時間的にオフセットされた比較プロセスが動作するフルタイム ウィンドウの観点からは意味があります。具体的には、以前の予測信号を約 50 ~ 100 ミリ秒後に到着する感覚信号と比較する必要があり、特定の運動動作に関連する感覚信号のウィンドウが 500 ミリ秒以上に広がる可能性があるため、進行中の比較プロセス ([[#figure_env]]) の時間エンベロープ全体が合計 1 秒以上に及ぶ可能性があります。したがって、おそらく 1 Hz のベースライン発火速度は、比較プロセスのこの動作エンベロープを反映しています。この動作エンベロープは 1 秒ごとにリセットされ、最初の下降動作信号と IO 糸球体内のギャップ結合接続によって同期されます。
IO 主導の学習ルール
小脳の学習ルールを定義する最後のステップは、[[#eq_delta-w-t]] を、IO エラー信号がある場合、およびシナプス減衰を引き起こすベースライン スパイクがある場合に機能する方程式に変換することです。
{id=”eq_delta-w-io” title=”IO 主導の学習”} \(\デルタ w = \lambda_e x_{t-\epsilon} \; \; \rm{エラー}\)
\[\デルタ w = -\lambda_d x_{t-\epsilon} \; \; \rm{衰退}\]ここで、$\lambda_e$ はエラー試行の学習率、$\lambda_b$ は減衰学習率です。減衰学習は、一般的にすべてを減衰させるのではなく、減衰学習をトリガーするベースライン 1 Hz スパイクの時点でアクティブな入力に対して動作することに注意してください。 PCDn 学習の場合は符号が反転します。
この学習ルールの非対称な方向性を裏付けるように、そのような強い非対称性を示すケースが複数のパラダイムにわたって十分に文書化されています。たとえば、VOR ([[#vestibulo-ocular reflex]]) パラダイムの非常に徹底的な研究では、VOR ゲインの増加のみが迅速なトライアルごとの調整を示し、これは PC クライミング ファイバー シナプス ([[@KimpoRinaldiKimEtAl14]]) の LTD と一致しました。それにもかかわらず、行動と PC の発火における長期的な変化は適切な双方向性であり、非対称性が急速で一時的な変化に限定されていることが示唆されました。この非対称性には、計算モデルを使用して調査できる一般的な機能上の利点がある可能性があります。
方向性サッカード課題も、急速学習において同様の非対称性を示しましたが、これも長期学習における双方向の可塑性とは切り離されていました ([[@YangLisberger14a]])。広く研究されているまばたき条件付けタスクでは、学習の方向性に関してより曖昧な結果が示されています ([[@McCormickThompson84a]]; [[@BerthierMoore86]]; [[@JirenhedBengtssonHesslow07]]; [[@GreenSteinmetz05]])。これはおそらく、学習の順方向モデルと適応フィルター モードの両方の組み合わせと一致しています。
また、苔状線維入力の性質の変化や、PC ニューロンを阻害する分子層介在ニューロンの可塑性など、PC 単純スパイク応答全体の可塑性を促進する要因が多数確認されています ([[@BonnanZhangGaffieldEtAl23]]; [[@ZhangYangGaffieldEtAl23]])。これらの要因の一部は PC 学習を調整する可能性があり、通常の 1 Hz バックグラウンド スパイク ([[@ZangDeSchutter19]]、[[@GuoUusisaari25]]) ではなく、意味のある IO スパイクに応答してのみ発生します。
標準モデル
長年にわたり、小脳機能については多数のさまざまな理論的説明がなされてきました (最近の総説については、[[@NguyenPerson25]]、[[@Shadmehr20]]、[[@RaymondMedina18]] を参照)。これらのモデルのほとんどは小脳皮質の回路に焦点を当てており、CN の回路には比較的ほとんど注意が払われていません ([[@KebschullCasoniConsalezEtAl24]]; [[@HerzfeldHallTringidesEtAl20]])。このセクションでは、上で示した核の枠組みに関連して、さまざまな理論間の相違点のいくつかの重要な点を検討します。
広範なレベルでは、Nuclear モデルによってサポートされる適応フィルタリングと順方向モデルの機能は、長年広く議論されてきた小脳機能の原則 ([[@Fujita82a]]; [[@MiallWolpert96]]; [[@Bastian06]]; [[@DeanPorrillEkerotEtAl10]]; [[@TanakaIshikawaLeeEtAl20]]; [[@Cullen23]]) と一致しています。それにもかかわらず、核モデルは、IO における予測誤差信号の起源と性質、および以前の説明では完全には統合されていなかった小脳核内のさまざまなニューロンの役割について、生物学的に特異的な機構的な説明を提供します。
[[@^DeZeeuw21]] は、よく研究されているさまざまな領域におけるアップバウンド学習とダウンバウンド学習の例を示し、関与する抑制性シナプスの数が方向性 (奇数 = アップバウンド、偶数 = 下バウンド) を決定すると主張しています。ただし、彼はこの区別を適応フィルタリングと順方向モデルのフレームワークに直接結び付けていません。さらに、初期の PC ベースのアップバウンド学習とダウンバウンド学習の統合を可能にするために必要な詳細な CN 回路は、[[@^DeZeeuw21]] 提案では特に取り上げられていませんでした。この機構は、上向きのマイクロゾーンにおける MDJ (中脳接合部) の独特の役割と合わせて、[[@^DeZeeuw21]] によって強調される出力極性の違いをはるかに超えています。
興味深いことに、最近の研究では、上向きと下向きのマイクロゾーンの強力な空間構成はげっ歯類に特有である可能性があり、霊長類ではより混合された構造([[@HaoZhuHuangEtAl24]])が示されており、これによりこれらの種間のさまざまな違いが説明される可能性があります(たとえば、サルの[[@KimpoRinaldiKimEtAl14]]とマウスの[[@VogesWuPostEtAl17]]を比較)。
おそらく、文献に残る不確実性と全体的な混乱の最大の原因は、より柔軟で制限のない運動制御における小脳の役割に関するものです。多くの人は、小脳がこの機能の中核部分をすべて単独で管理できると考えており、その場合、かなり複雑で困難な計算問題を解決する必要があります (例: [[@Kawato99]]、[[@Llinas09]])。しかし、多くの証拠は、[[motor]] ページでレビューされているように、そのような能力には [[basal ganglia]] と [[prefrontal cortex]] を含む複数の脳領域間の調整された相互作用が必要であることを強く示唆しています。したがって、小脳を、それぞれが明確に定義されたタイプの神経計算と学習に特化した、さまざまな神経システムのより広範な文脈に置くことで、おそらく、より満足のいく全体的な理解に到達できるでしょう。
IO エラー信号の性質
興味深いのは、小脳機能に関するほとんどの議論では IO が誤差計算を実行していると描写されており、多くの論文では予測誤差コーディングとこれらの詳細な生物学的特性の両方が議論されているにもかかわらず ([[@LoyolaHooglandHoedemakerEtAl23]]; [[@DeZeeuwSimpsonHoogenraadEtAl98]])、IO ニューロンの固有の生物学的特性とこの誤差計算との間の直接的かつ明示的な関係を明確に示した論文はありません。
[[@^OhmaeMedina15]] によって提供される IO エラー信号の概要と、多くのまばたき条件付け研究にわたる学習は、IO ([[@Hull20]]; [[@NguyenPerson25]]) での新しい形式のエラー計算を示すものとして強調されています。ただし、これらのプロパティはすべて、CNiIO 予測学習経路 ([[#eq_delta-w]]) に対して仮定されたデルタ ルール学習アルゴリズムと一致しています。この方程式は、レスコルラ・ワグナー学習則 ([[@RescorlaWagner72]]) としても知られており、もともとは、一時的なドーパミン発火に関連する広範囲の条件付け現象を説明するために使用されていました (詳細については、[[PVLV]] を参照してください)。この学習で潜在的に困難な唯一の側面は CS ベースの応答ですが、これは小脳予測学習の予期バイアスと一致しています。
{id=”figure_raymond-medina” style=”高さ:25em”}
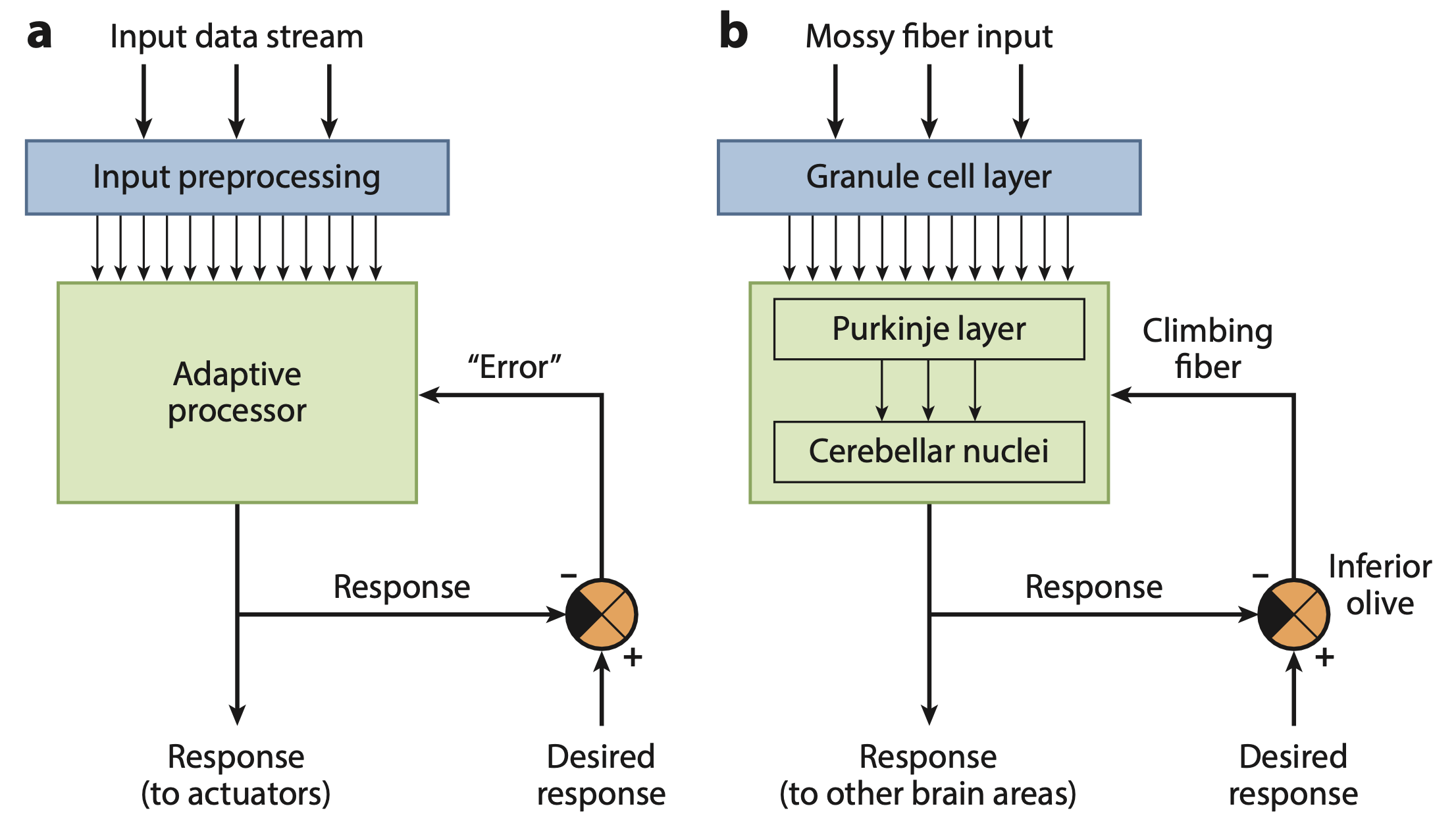
[[@^RaymondMedina18]] から [[#figure_raymond-medina]] に示されているように、感覚予測誤差を計算するための核モデルの明確で生物学的に明示的なメカニズムとは対照的に、小脳機能のこの重要な側面の他のほとんどの処理は著しく曖昧です。脳は「望ましい反応」のような全知のものをどこでどのように計算するのでしょうか?そして、そのようなものが利用できるのであれば、最初から望ましくない応答をするのではなく、それを直接使用してみてはいかがでしょうか?
実際、エラー信号を直接計算可能にするために必要なステップは、感覚結果の観点からエラー信号を定義することです。これは、これらが小脳に投射される具体的な結果信号であるためです。しかし、小脳は運動制御に関与しているため、人は当然、感覚エラーではなく運動エラーの観点から考えたくなります。ここで問題が発生します。次に説明するように、「モーターエラー」をどのように定義するかが非常に不明確です。
運動空間は乱雑だが、感覚はすっきり
| モーター制御、およびモーター制御空間での表現に関する考え方の根本的な問題は、それが非常に厄介な場所であるということです。具体的には、運動動作の望ましい結果と、その結果を達成するために必要な実際の筋肉の動きの間には、一般にそのような動作ごとに異なる膨大な数の変数に応じて、大規模な 1 対多の関係が存在します。テーブルの上のカップに手を伸ばすことを考えてみましょう。この単純な動作を成功させるために必要な正確な運動動作は、腕の各関節の初期位置、体の残りの部分の姿勢構成 (立ったり座ったり、バランスなどの要素との正確な関係)、カップの物理的特性、筋肉の疲労状態などに依存します。これらの各要素が相互作用し、大規模な [[curse of dimensionality | combinatorial explosion]] 問題を引き起こします。 |
このため、感覚空間 ([[@Powers73]]) でモーター制御を駆動する方がはるかに簡単です。感覚空間では、運動動作の目標とその結果の両方がすべて、はるかに低次元の、より体系的な表現空間にエンコードされます。したがって、カップに手を伸ばすと、視覚、体性感覚、固有受容の信号によって、その動作が成功したかどうか、どの方向に失敗したかなどがわかります。
さらに、適応フィルタリング プロセスは感覚空間を体系的にクリーンアップできるため、上記のすべての要素 (開始関節角度、姿勢、バランスなど) が関連する感覚信号からすでに差し引かれています。したがって、このフィルタリングされた感覚空間に基づく運動計画は、より体系的で低次元になります。
この感覚ベースのフレームワークは、順モデルを反転して「逆モデル」 ([[@JordanRumelhart92]]; [[@Kawato99]]) を取得する試みとは対照的です。逆モデルは、原理的に、特定の感覚結果を得るためにどのような運動コマンドが必要かを示します (順モデルが逆に進み、運動コマンドの感覚結果を予測していることを思い出してください)。上で説明したすべての理由により、同じまたは類似の結果をもたらす可能性のあるアクション シーケンスが多数存在することが多いため、複雑な現実世界のケースで逆モデルを作成することは非常に困難です ([[@JordanRumelhart92]]; [[@GibsonFichmanPlaut97]])。
モーターベースのフレームワークの問題についてのさらなる洞察は、遠心性コピー入力に基づくことが提案されている、IO によって計算される予測誤差信号の考察から得られます ([[@Oscasson80]]; [[@KawatoGomi92]])。 [[@^SimpsonWylieZeeuw96]] は、入力が遠心性コピー入力ではなく、体性感覚信号と固有受容信号によって支配される傾向があることと、最初の運動コマンドの遠心性コピーとその後の運動結果との間の時間的分離が比較的長い可能性がある ([[#figure_env]] に示すように 100 ミリ秒以上) 可能性がある、運動誤差の計算にタイミングの問題があることに注意してください。さらに、下行運動経路と上行運動経路は小脳にマッピングされるため、解剖学的に不一致があります ([[@DeZeeuwSimpsonHoogenraadEtAl98]])。
経路はモデルに含まれていません
以下の経路は文書化されていますが、現在のモデルには含まれていません (一般に、これらの経路は広く議論されていません)。
- [[@^UusisaariKnopfel10]] は、CN 内のグリシン作動性 (抑制性) 非緊張活性ニューロンからの小脳皮質への投射を実証しました。これらは比較的まばらで、晩生でした。
## まとめ
要約すると、感覚予測学習 (順方向モデル) と適応フィルタリングは、Nuclear モデルで捉えられている既知の小脳特性と一致する、完全に計算的に明確に定義されたフレームワークを提供します。この適応フィルタリング機能だけを使用して機能的なモーター制御を実装し、運動動作の基礎となる高品質の感覚信号を提供することができれば、多くの困難な問題は解消されます。 [[motor]] ページで提供される広範なフレームワークと、[[basal ganglia]] および [[prefrontal cortex]] の機能を含む実装された計算モデルは、これが実際に可能であることを示しています。
さらに、[[Rubicon]] 目標駆動制御フレームワークは、望ましい目標結果を達成するための運動動作選択のための重要なアウターループ インフラストラクチャを提供します。このシステムは、複数レベルの予測学習システムによって通知され、学習された [[distributed representations]] にわたる [[constraint satisfaction]] 処理によって形成される感覚ベースの目標選択メカニズムを使用して、望ましい結果と全体的な行動計画を選択します。この目標選択プロセスは、小脳の予測学習メカニズムによって情報を得ることができますが、逆モデルを取得するために順モデルを反転する必要はありません。
前庭眼球反射
{id=”figure_vor-anatomy” style=”高さ:30em”}
![VOR 反射の解剖学。VOR 反射は、三半規管からの前庭信号によって駆動され、前庭核複合体 (VNC) に投射され、そこから光学運動野に投射されます。すべての一次入力は同側性 (例: 右耳が右半球に投影される) であり、VOR には、代償的な反対方向の眼球運動を引き起こす _外転核_ 光学運動介在ニューロンを駆動するために、_対側性_ に投影する VNC ニューロンのサブセットが含まれます。反対側の経路によって阻害される同側経路や、半球を越える阻害性競合も存在します。これは典型的な対戦相手プロセス組織です。この回路の VNC ニューロンは、他の領域の小脳核ニューロンと機能的に同等であるため、これらのニューロンの直接的な可塑性が学習の最終的な統合結果となります。この反射における利得の小脳皮質の調節は、プルキンエニューロンからVNCニューロンへの強直性神経活動の変化(例えば、脱抑制により利得が増加する)、およびこれらのVNCニューロンへのシナプス入力の変化から生じる。 [wikimedia.org] から転載](https://commons.wikimedia.org/wiki/File:Vestibulo-ocular_reflex_EN.svg) .] (media/fig_cerebellum_vor_anatomy.png)
.] (media/fig_cerebellum_vor_anatomy.png)
{id=”figure_vor” style=”高さ:30em”}
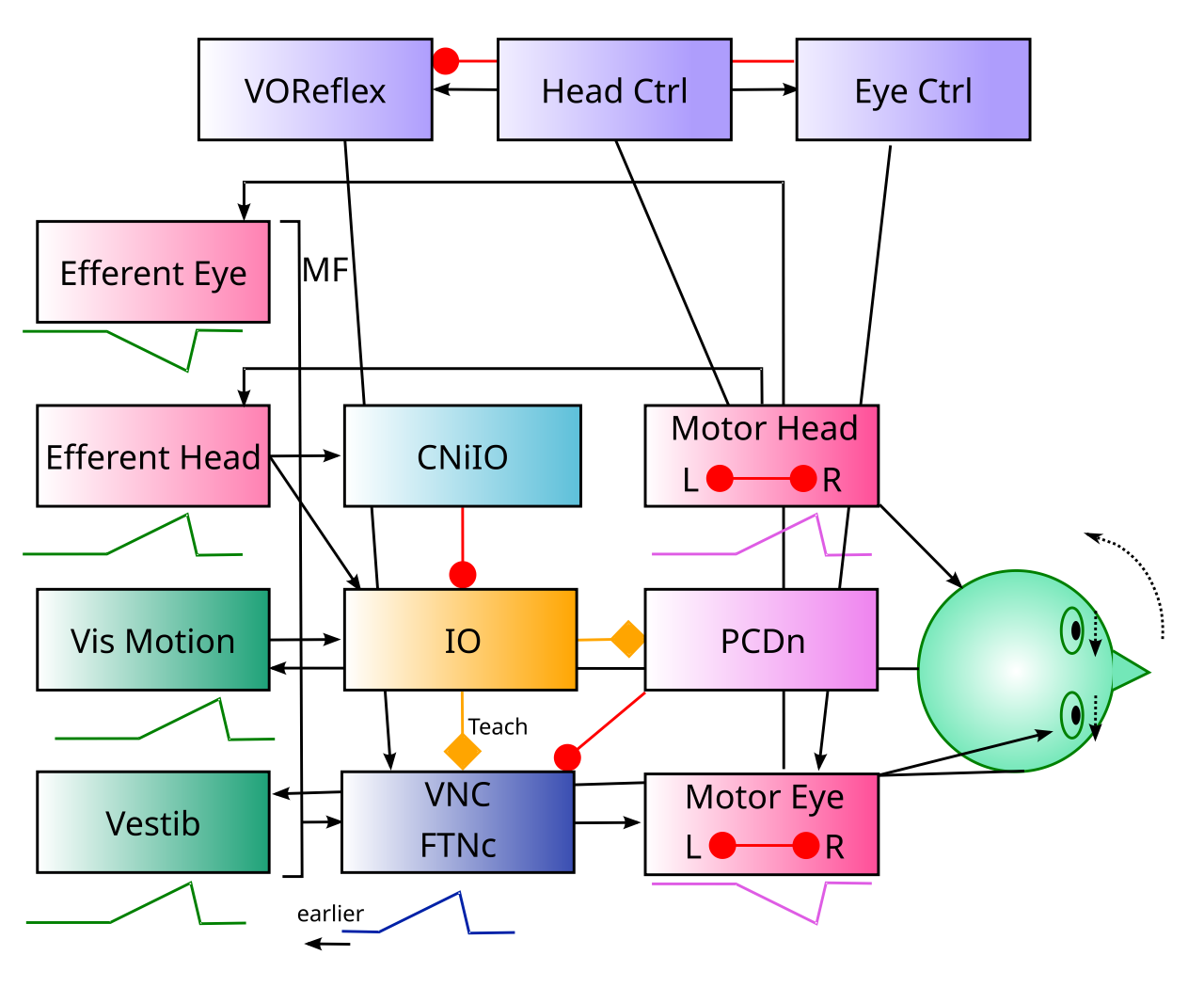
前庭動眼反射 (VOR) は、小脳機能の徹底的に研究された側面です (レビューについては [[@Cullen23]]、[[@KheradmandZee11]]、重要な結果については [[@ZeeuwHanselBianEtAl98]]、[[@KimpoRinaldiKimEtAl14]] を参照してください)。これは、頭の動きが始まると、反対方向に動くことでその頭の動きを補うために予期的な眼球運動が引き起こされる、前方モデル、つまり小脳機能の下向きの側面のよく理解された例を提供します。最終的な結果は、網膜上で比較的安定した画像となるはずであり、全体的な視覚運動信号 (つまり、網膜滑り) が IO エラー信号を駆動し、それが小脳の学習を駆動します。
VOR は、[[vestibular]] 感覚入力 ([[#figure_vor-anatomy]]) によって駆動される脳幹回路によって直接仲介されます。小脳機能の進化上最も古い前庭系の側面のこの場合、図に示すように、小脳核ニューロンの機能的等価物は二次前庭核複合体 (VNC) 内に直接位置しています。これらのニューロンは、VOR 反射に直接関与しますが、他の場合では、小脳核ニューロンは通常、MDJ や他の領域など、他の脳幹回路を調節します。
[[#figure_vor-anatomy]] に示されている反射回路は、脳幹のさまざまな部分にある多くの同様の回路の典型であり、反射を有効または無効にするために調節できる二次ニューロンを介して、感覚信号が関連する運動行動を直接駆動します。
これは反射として説明されていますが、VOR に対する主な小脳の寄与は、頭の動きの感覚効果を (下向きの前方モデル経路を介して) 予測することです。そのため、代償性の眼球運動は必ずしも遅れているわけではなく、むしろ先取りして頭の動きの影響をより正確に打ち消すことができます。
論理的には、この順方向モデルは、頭の動きによって生成される網膜からの実際のフルフィールド視覚運動信号に基づくものになります。なぜなら、これらの視覚運動信号には、キャンセルされる実際の運動信号に関連する時間エンベロープがあるからです。ただし、これらの信号にはキャッチ 22 が関連付けられています。代償性の目の動きによって信号はゼロになります。したがって、予期される特定の運転感覚入力が前庭感覚から得られることが重要であり、アクティブな VOR 眼球運動であっても頭の動きによって常に活性化されます。
実験研究では、小脳の寄与は、VOR ゲイン係数、つまり頭部の動きと反対方向の代償性眼球運動の比率への寄与によって要約されます。正常なベースラインの場合、この利得係数は 1 です。しかし、(眼鏡を介して)目の倍率を変更したり、前庭神経を刺激したりするなどの実験的操作により、小脳に依存して利得が変化します。
VOR ゲインは乗算係数ですが、一般にニューロンは他のニューロンに乗算効果を直接引き起こすことはできません。代わりに、これらの回路でこれを実現する方法は、[[opponent]] 組織を通じて行われるため、調節ニューロンはターゲット動作に向かって引っ張る動作と、ターゲット動作から遠ざける動作の両方を興奮させます。興奮性ドライブのレベルが調節されると、これらの敵対するニューロンの発火の全体的な範囲も変化し、この全体的な範囲が有効なゲイン係数に対応します。
TODO: エラー信号としての視覚的な動きがゼロであることが学習にどのように変換されるかという重要な問題 – このゼロ化は、おそらく他の多くの場合にも当てはまります。
生物学に基づいたモデルで考慮すべき関連現象には、次のようなものがあります。
-
学習率と保持期間の観点から、VOR ゲインの非対称性は増加するか減少します ([[@BoydenKatohRaymond04]]; [[@KukiHirataBlazquezEtAl04]]; [[@KimpoRinaldiKimEtAl14]])。これらの違いは、ゲイン増加に対する下向き小脳皮質 LTD 学習の急速な寄与(比較的急速に減衰する)と、有効小脳核ニューロンにおけるゆっくりとした長期にわたる変化([[@LisbergerPavelkoBroussard94]])の観点から理解できます。
-
[[@BlazquezHirataHeineyEtAl03]] は、さまざまな入力パラメータに対するプルキンエ細胞の応答におけるさまざまな変化を文書化しており、これらを合わせて VOR ゲインの変化を説明できます。
-
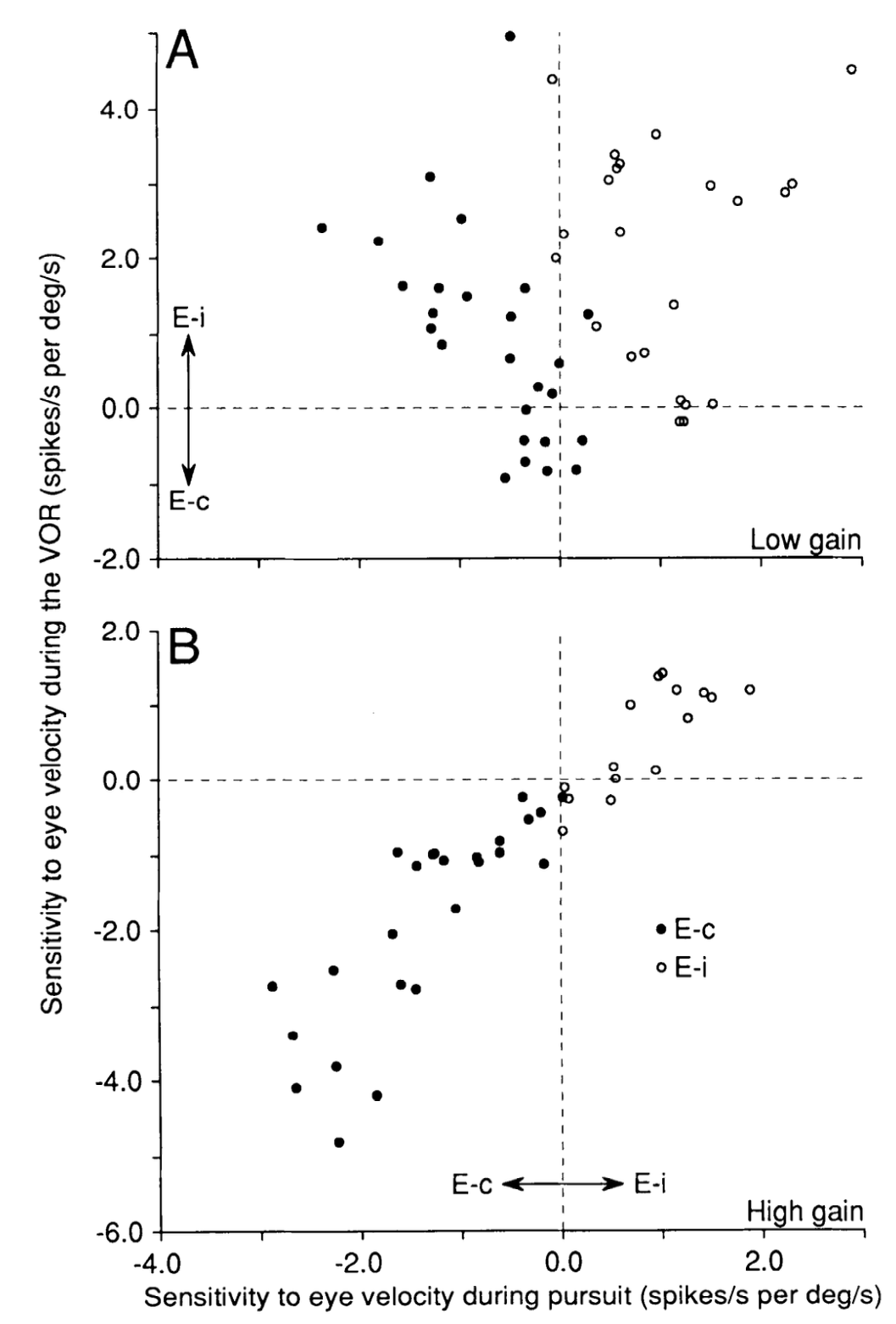
[[@^LisbergerPavelkoBroussard94]] (例: [[#figure_vor-ftn]]) で文書化されている、同側性対対側性発火特性を持つものを含む、綿状標的ニューロン (FTN) として知られる、VOR を媒介する二次前庭ニューロンの変化の詳細なパターン。
{id=”figure_vor-ftn” style=”高さ:40em”}
既存のモデルのほとんどは比較的抽象的であり (例: [[@HollandSibindiGinzburgEtAl20]]、[[@InagakiHirata17]]、[[@ClopathBaduraZeeuwEtAl14]]、[[@Lisberger09]]、[[@KawatoGomi92]]、[[@Robinson81]])、プルキンエ関数の学習を促進する重要なトレーニング信号を IO が計算するための詳細なメカニズムを提供するものはないようです。そして前庭核細胞。ただし、その重要な違いを除けば、[[VOR simulation]] で調査できる Nuclear モデルは、これらの既存のモデルによって確立された原則と全体的に一致しています。