compcogneuro/web: evolution
このページは外部資料の日本語訳です。原文の見出し順と本文順を保ち、コード・URL・出典表記はできるだけ原形のまま残しています。
出典とライセンス
原典: https://github.com/compcogneuro/web/blob/main/content/evolution.md
ライセンス: Text: CC BY 4.0; code: BSD 3-Clause。このページは日本語翻訳であり、変更点は翻訳とサイト内整形です。
+++ Categories = [“Neuroscience”] bibfile = “ccnlab.json” +++ 進化のプロセスは、事実上、DNA にコード化された発生プロセスに対する大規模並列 [[search]] アルゴリズム (つまり、[[genetic algorithm]]) であり、自己複製能力を最適化します。資源が限られているという状況では、これは競争をもたらし、地球上の生命の長い血なまぐさい戦いは当然の結果です。
何百万年にもわたるこのプロセスの結果は、種の生存と複製を最適化するために哺乳類の脳が組織化される方法に反映されています。他の獣と比較して、哺乳類はより精巧な機能を備えた大きな脳を持っており、特にこのハードウェアなしでは不可能な認知能力を可能にする広範な [[bidirectional connectivity]] をサポートする [[neocortex]] を備えています。
[[Rubicon]] フレームワークは、努力やリスクなどのコストと比較したプラスの結果の全体的なバランスを最適化する方法で、時間の経過とともに行動を組織化できる効率的な目標主導型の行動を促進するために、複数の脳領域を組織化するために進化が発見した教訓を発見するという [[computational-cognitive-neuroscience#reverse engineer the brain]] への試みです。この仮説は、進化の並行探索プロセスは、個々のモデルの「寿命」内で動作する一般的なエラー駆動型の学習メカニズムでは容易に再現できないため、適切なニューラル システムによって理解および実装される必要がある、というものです。
{id=”figure_evol” style=”高さ:60em”}
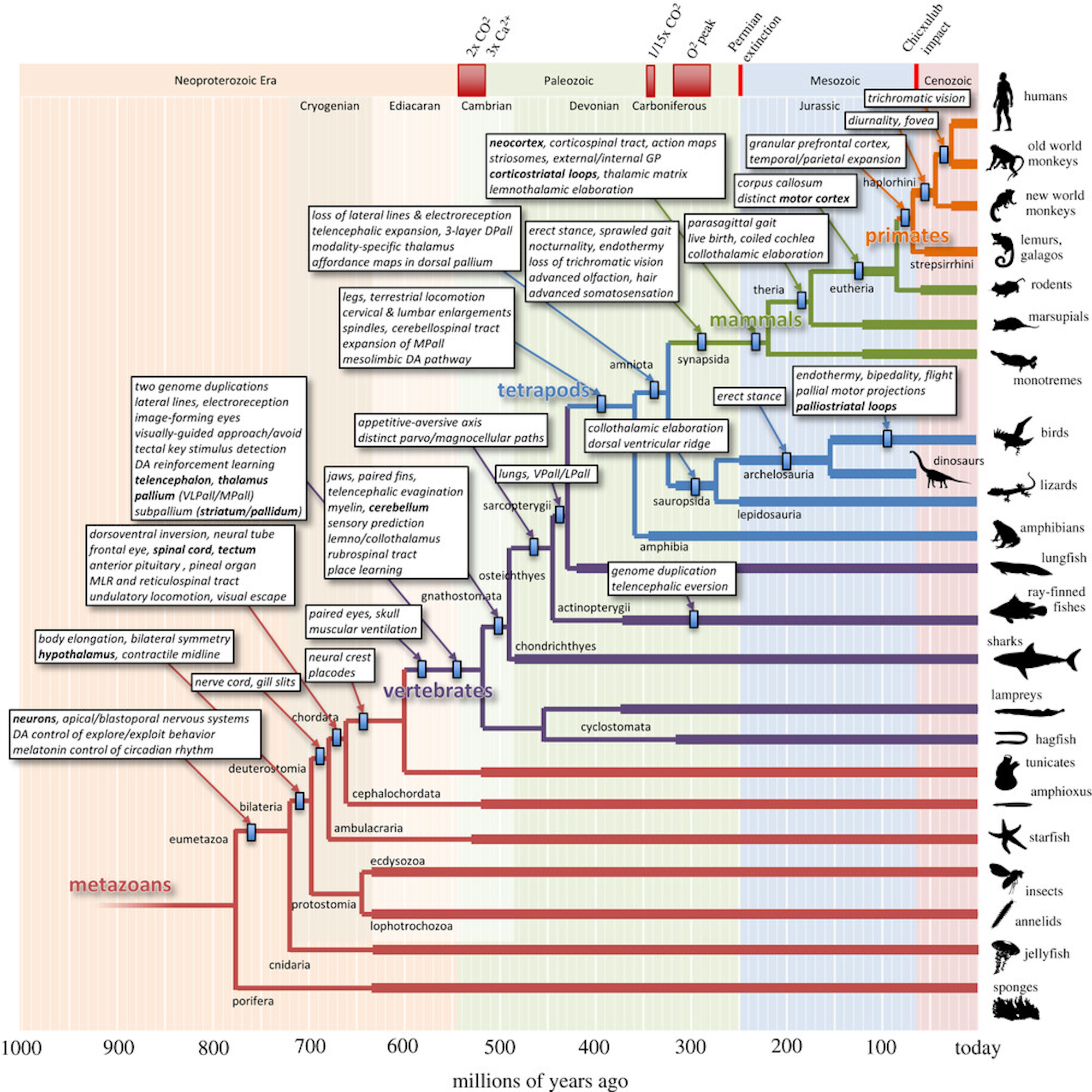
[[#figure_evol]] は、哺乳類、霊長類、そして最終的に人類に至る進化の軌跡に関連する脳構造の重要な変化の一部を記録する [[@^Cisek21]] の驚くべき図を示しています。これにより、時間の経過とともにさまざまな機能が追加されてきた方法の全体像がわかります。進化の軌跡には主に 2 つの「戦略」が見られます。
-
既存の構造を精緻化および拡張し、効果的なアルゴリズムを微調整することで構造をより大きく、より機能的にします。
-
新皮質などのまったく新しい神経アーキテクチャを使用して、既存の構造の上に新しい構造を追加します。この場合、既存のシステムは通常、[[subsumption]] アーキテクチャ ([[@Brooks86]]) に準拠して、新しいシステムによって変調制御されます。
{id=”figure_tube” style=”高さ:45em”}
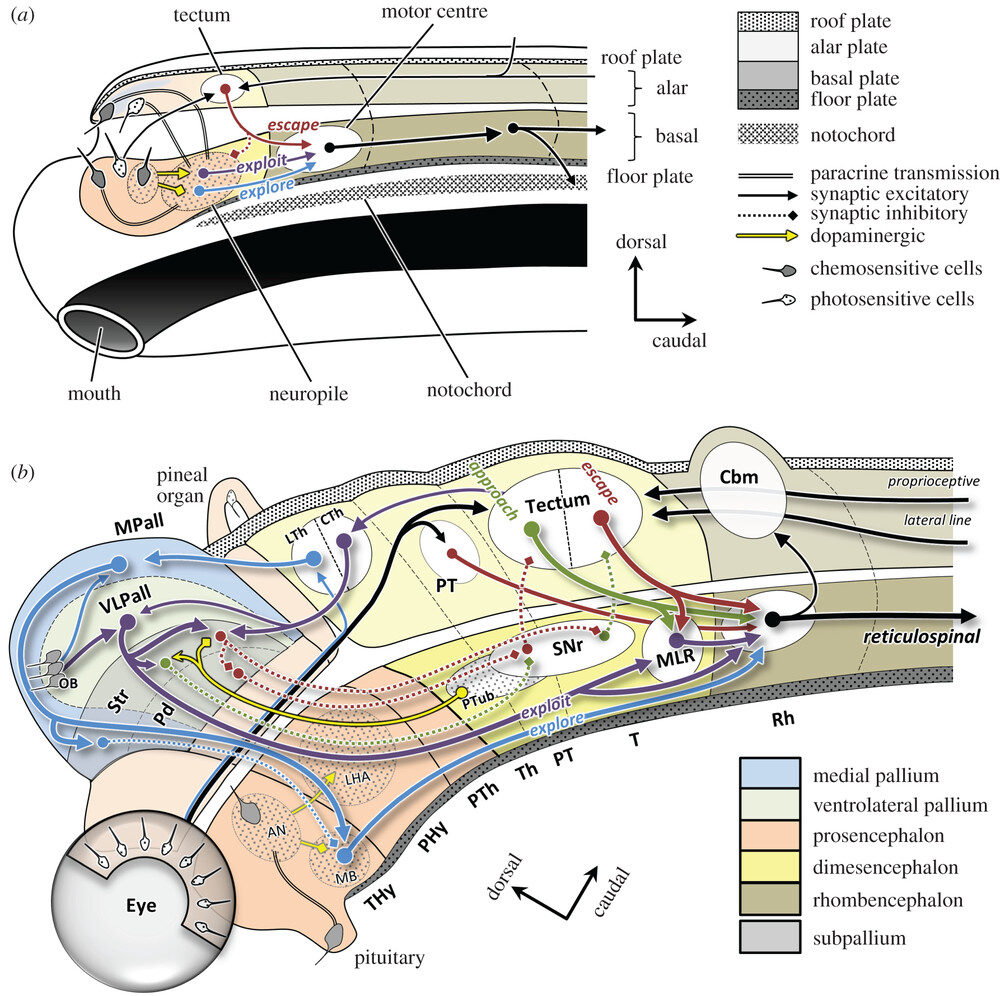
[[#figure_tube]] は、これら両方のプロセスの例を、ナメクジウオの脳 ([[#figure_evol]] ではヒトデのすぐ上) から初期の顎口類の顎のある魚 (サメが現代の例です) への変化において示しています。すべての脳の発達と構造のテンプレートを提供する既存のチューブ状の bauplan ([[@NieuwenhuysPuelles15]]) は、進化の軌跡を経て保存され、精緻化されています。
進化は主に DNA によって組織化された発生プログラムに基づいて行われるため、発生の軌跡は進化の軌跡を反映しており (つまり、個体発生は系統発生を再現します。; [[@Gould77]])、哺乳類であっても、すべての異なる脳部分が最終的にどこに到達するかが決まります ([[anatomy]] を参照)。したがって、背側_経路は主に感覚系であり、_腹側_経路は主に運動系です。すべてが中心核から放射状に広がるにつれて、_内側 領域が最初に発達し、そのため進化的により古いシステムを持つ傾向があります。したがって、生存に関連する [[emotion]]al および行動を駆動する動機付けシステムは、一般に腹側および内側にあり、背外側 [[prefrtontal cortex]] などの後期に発達するシステムは感覚および認知です。
脳は、実質的に管の前端で感覚経路と運動経路を結び付けており、[[hypothalamus]] に相当する「ナメクジウオ」に存在する最高レベルの脳領域で、摂食 (搾取)、採餌 (探索)、逃走という 5 つの F ([[@Pribram60]]) の基本的な運動動作を駆動する栄養素摂取の基本的な感知を提供します。 (逃亡)、喧嘩、そして淫行。
| 顎のある魚には、視床下部 (LHA) の上に構築され、体の動きの [[space | spatial]] 処理に重要な乳頭体 (MB)_ を含む多くの追加構造が存在します。 MB は_内側パリウム (MPall)_ と相互接続し、これが [[hippocampus]] になるものの始まりです。 [[cerebellum]] はここで、特にチューブの背側感覚部分に初めて登場し、魚の固有受容感覚と [[vestibular]] 感覚に相当する側線感覚入力を処理し、動物の自己運動の影響を予測してキャンセルする方法を学習し、説明のつかない摂動が明らかになるようにして、[[motor]] の適切な応答を駆動します。アクション。 |
[[basal ganglia]] のコア要素 (Str = 線条体、Pd = 淡蒼球、SNr = 黒質網様体) が存在し、BG の Go 経路と No 経路に対応する amphioxus から継承した approach 対 escape 回路のテンプレートが存在します。 パリウム (淡蒼球と混同しないでください) は、後に_テレンスフェロン_の [[neocortex]] となるものの始まりであり、腹外側パリウム (VLPall) は、嗅球 (OB) 化学感覚からの直接入力と視器官からの視覚入力を使用して、BG 作用システムのアウター ループのゴール駆動型調節を提供するように配置されています。 (哺乳類の [[superior colliculus]] の前駆体)。 BG の進化の歴史の詳細については、[[@^GrillnerRobertson16]] を参照してください。
驚くべきことに、5億年前の初期の顎のある魚類に存在する、進化上古代の脳テンプレートでさえ、哺乳類の脳に存在するのと同じ機能と組織の多くを備えており、これらの単純な脳システムを理解することは、より複雑な脳システムの機能と組織を理解するのに役立ちます。
精緻化と付加という進化のプロセスの観点から見ると、この強固な機能基盤が確立されていれば、海馬、大脳基底核、小脳などの哺乳類のバージョンへの原要素の相当量の精緻化が可能になります。新皮質のルーツはパリウムにありますが、それはより多くの新しい機能の追加を表します。
進化のプロセスのもう 1 つの意味は、生存に関連する中核的な機能が既存のニューラル アーキテクチャによって処理されると、新しい開発では特定の能力を最適化するための「自由」が増えるということです。これは、新皮質系と皮質下系の区別と一致しています。新皮質系は通常、特定の行動に対して遺伝的に強く制約されているのに対し、新皮質は柔軟な学習能力を自由に最適化できます。
したがって、多くの皮質下領域は、同じ回路内ですべて絡み合った複雑で一見恣意的な機能の組み合わせを持っています。たとえば、小脳の断端核と虫歯、腹側内側線条体(特に側坐核の殻)、扁桃体の中心核、視床下部全体などです。対照的に、新皮質には、比較的一貫した学習表現を備えた広い領域があります。基本的には旧市街と郊外の違いです。
メカニズム
進化論は一見すると不合理な理論です。魚のような獣が魚ではないものを生み出すことなど、ほんのわずかでもあり得るのでしょうか?そして、たとえできたとしても、その「変人」はどうやって他のものと交尾して、その不自然さを広めることができるでしょうか?この新種を繁殖させるのに十分な距離に、2 匹の奇妙な非魚類が同時に出現したのだろうか?それはまったく意味がありません。間違いなく、この基本的なレベルでの考え方のまったくの信じがたいことが、人々がこの理論に対して持ち続ける抵抗感の一因となっています。
{id=”figure_cells” style=”高さ:10em”}
{id=”figure_lego” style=”高さ:20em”}

進化がどのように起こり得るかを理解するための最初のステップは、動物は多数の微細な構成要素 (細胞) が組み立てられた結果であり、細胞レベルでは、実際には魚と人間の間に大きな違いはないことを理解することです ([[#figure_cells]])。したがって、同じレゴ ブロック ([[#figure_lego]]) のセットから、巨視的に異なる多くのものを構築できるのと同じように、生物学では発生プロセスを微調整するだけで、非常に異なる見た目の種類の獣を比較的簡単に構築できます。この発達プロセスは、レンガの組み立て方を説明するレゴの小冊子の指示に従うプロセスに似ており、小冊子は DNA に似ています。
2 番目の重要なステップは、発生プロセスが実際に進化がいじる主なものであり、そうすることによって、たとえ非常に小さな遺伝的変化であっても巨視的な外観を劇的に変えることができることを理解することです。 LEGO の例えを続けると、説明書には N 回繰り返す手順が含まれることがよくあります。N の値を変更すると (説明の長いリストの 1 つの数字にすぎません)、結果として得られるコンポーネントのサイズを簡単に変更できます。同様に、進化は、特定の発達段階を繰り返す回数を比較的簡単に変更し、結果として生じる脳領域や身体部分のサイズを劇的に変えることができます。たとえば、ジョセフ・メリック (Elephant Man、wikipedia リンクとして知られています) には、発達プロセスの比較的遅い段階で出現した広範囲にわたる成長が見られました。
発達のプロセスは非常に堅牢で、指示に柔軟に適応できます。これは、片方の親からの遺伝的変化が依然として発達の結果を形成する理由を説明するのに役立ちます。両方の親を調整して変更する必要はありません。これらの変化が生存に有益であれば、それは世代を超えて受け継がれます。
最後の大きな課題は、どれほど劇的な「質的」な変化が起こり得るかを理解することです。たとえば、魚はエラから肺へと、どのようにして陸に移動することができたのでしょうか?まず、これは満潮時に座礁する可能性が十分にある潮だまりのような環境で発生した可能性があります。第二に、肺は当初、閉じ込められた空気を利用して浮力を制御するメカニズムによって発達した可能性があります。この閉じ込められた空気を呼吸に利用できる魚は、それができない魚よりも多くのエネルギーを得ることができたでしょう。これは、潮汐環境下での生存に明らかな利点をもたらすでしょう。
同様のプロセスが鳥の羽にも起こり、最初は温度調節のために開発され、その後飛行に役立つようになりました。 Stephen J. Gould はこのプロセスを「exaptation」 ([[@GouldLewontin79]]) と呼び、ダーウィンはこれを「preadaptation」と呼びました (これはグールドにとっては少し「先見的」すぎるように聞こえました)。重要な点は、遺伝子検索プロセスには多くのノイズがあり、あることに少し役に立ったものが、別のことに非常に役立つ可能性があるということです。
驚くべき事実は、あなた方から何十億年も遡る「存在の偉大な連鎖」が実際に存在し、各世代が生存のために戦い、ほとんど非常に小さな方法で革新し、時折劇的な進歩を遂げているということです。そして、[[#figure_cells]] をもう一度見てみると、このレベルでは、この数年間で実際にはそれほど変わっていないことがわかります。
大脳新皮質の進化
[[#figure_tube]] に示されている bauplan は、脊椎動物の脳全体の多くの異なるコンポーネントがどのように組み合わされているかについての全体像を理解するのに役立ちます。哺乳類の強力な認知能力に不可欠な [[neocortex]] は、進化の観点から、この全体像にどのように適合するのでしょうか?
第一に、大脳新皮質のユニークな貢献は、運転行動の「より賢い」、より抽象的で体系的な基礎を提供する、新しい高次の感覚運動表現を「学習」する能力であるということで広く同意されています(例:[[categorization]])。これにより、蓋内の遺伝的にコード化された特徴検出器を超えて、生物の適応能力が大幅に拡張されます ([[superior colliculus]])。
{id=”figure_pallium” style=”高さ:25em”}
![(b) の羊膜類 (哺乳類、鳥類、爬虫類) と (a) の他の非羊膜脊椎動物 (魚類、両生類) との間のパリウムの漸進的な分化。 VLPall (腹外側外套) におけるより近位の刺激 (接近 vs 回避) の表現と、MPall (内側外套) におけるより遠位のナビゲーションに関連する刺激の表現の間には、最初の基本的な区別があります。 CTh = 丘によって駆動される冠視床、LTh = 直接網膜入力を受け取るレムノ視床。 VLPallは羊膜類でさらに腹側、側方、背側の外套に分化し、背側(DPall)は直接レムノ視床からの入力を受け取り、後に新皮質に変わります。 [[@Cisek21]] より。](https://raw.githubusercontent.com/compcogneuro/web/main/content/media/fig_evolution_pallium_cisek_21.png)
[[@^Cisek21]] は、新皮質が出現するパリウムの進化的分化に関する幅広い文献の中に新皮質の進化を位置づけています ([[#figure_pallium]])。たとえば、彼は、接近および回避行動を支配するより近位の世界の表現 (腹側外側外套、VLPall) と、ナビゲーションに重要なより遠位の世界の表現、内側外套 (MPall) の間には、進化的に古くから存在する区別があると主張しています。彼はさらに、この広範な区別は、この枝が進化した水生種と比較して、陸上の生活に適応するにつれて、哺乳類、鳥類、爬虫類(つまり、羊膜を持つ羊膜類)に至る過程でより複雑になり、分化していくと主張しています。
特に、脊椎動物が陸上を移動するにつれて、視覚は長距離のナビゲーション処理を推進できるようになり、海馬および関連領域の構造への内側外套の精緻化をサポートしました。パリウムの腹側と外側の領域も分化しており、特に新皮質は、より複雑な感覚運動アフォーダンスを学習するために重要なパリウムの背側領域から成長したと彼は主張しています。
この一般的なストーリーは、進化の歴史への重要な手がかりを提供する発生および解剖学的データに根ざしている可能性があります。具体的には、より最近の新皮質を取り囲む、進化と発達の点でより古い中皮質の環があり、これらの中皮質領域の性質と組織は、哺乳類の新皮質の進化の軌跡の説得力のある説明を提供します ([[@PuellesAlonsoGarcia-CaleroEtAl19]]; [[@Cisek21]]; [[@NordPattabiramanViselEtAl15]]; [[@BarbasZikopoulos25]])。この中皮質リングは多くの点で古典的な [[limbic system]] 構造に対応しており、「辺縁系」という用語は複数の場所で存続します。
{id=”figure_ring1” style=”高さ:35em”}
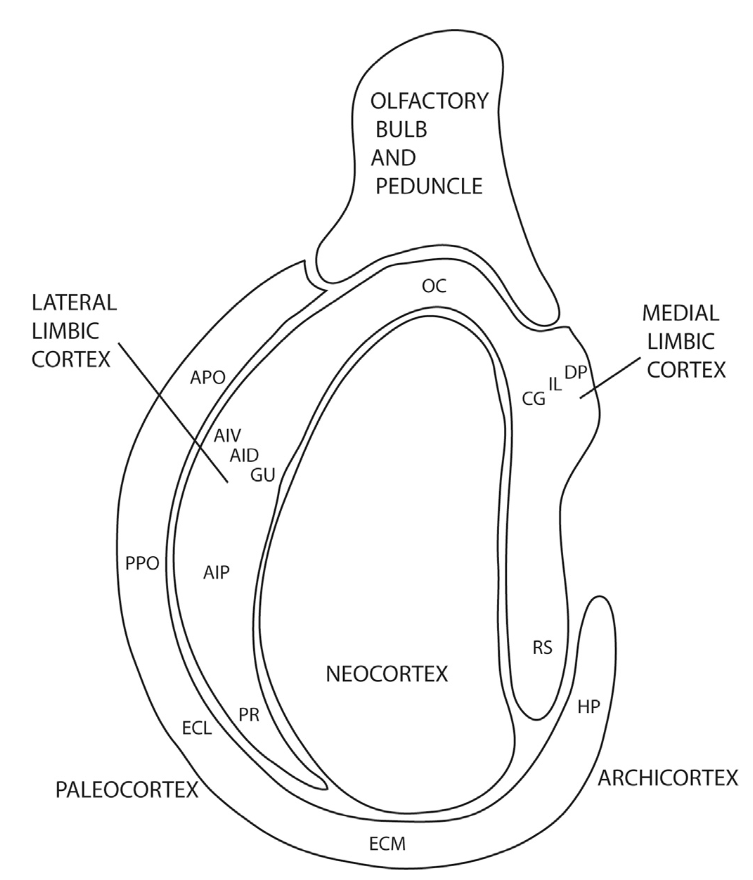
[[#figure_ring1]] は、以前の提案 ([[@Swanson87]]; [[@BayerAltman91]]) に基づいた、中皮質リング構造の簡易バージョンを示しています。 [[@Cisek21]] との核となる区別と一致して、リングを「前方」系と「後方」系に分割することで、2 つの主要な機能コンポーネントを特定できます。 [[Rubicon]] 目標駆動型フレームワークの観点から、前方システムを、望ましい結果を獲得し、否定的な脅威を回避するために、アプローチと回避行動を組織化するために必要な、より高いレベルの表現のすべてをサポートしていると解釈します。
具体的には、眼窩前頭皮質 (図の OFC、OC) は、特定の可能な結果の値を表します。これは、[[insula]] (AIV、AID) および味覚皮質 (GU) の外側辺縁皮質領域でより具体的に表されます。前帯状皮質(図のACC、CG)、辺縁下皮質(IL)、および辺縁前皮質(図のPL、DP)は、それぞれ、行動のコスト、より抽象的なレベルでの価値、および全体的な統合利益、つまりコストユーティリティを表します。
これらの皮質領域における生存に関連する結果のコード化は、たとえば OFC と [[amygdala]] の間など、関連する皮質下領域との広範な双方向接続を介して確立され、OFC が扁桃体で表される価値を満載した結果の予測を維持できるようになります。したがって、リング周囲の他の中皮質領域と一致して、OFC は新皮質の残りの部分と関連する皮質下システムとの間のゲートウェイを提供します。
これらの前中皮質領域はげっ歯類の脳の [[prefrontal cortex]] (PFC) 全体を構成しており (同ページの [[prefrontal cortex#figure_pfc-comp]] を参照)、これらの領域の視床皮質回路は、これらの重要な PFC 領域を欠いている非哺乳類の脳よりも、目標に基づいた積極的な計画を長期間維持する独自の能力を提供している可能性があります。
さらに、[[basal ganglia]] (腹側_pallium_と混同しないでください) の出力核である腹側淡蒼球 (VP) は、[[Rubicon]] モデルによれば、これらの PFC 領域に重要なゴールエンゲージメント_ゲート_ダイナミクスを提供します。進化および発生の軌跡は、大脳基底核と新皮質の間の強力な相互接続を示しており、新皮質はますます精巧な「状態空間」入力を大脳基底核の [[reinforcement learning]] 機構に提供しています。
また、この PFC ベースの視床皮質回路は、[[thalamus]] の中背核 (MD) に関与しており、[[predictive learning]] の中核機構をサポートしており、この機構は、高次の新皮質領域と相互接続する、後期に発達する視床の肺核によってもサポートされています。
| したがって、進化の軌跡は、回避行動ではなくより柔軟な生存主導のアプローチをサポートする、この中核となる作業記憶と予測学習機能が、新皮質の進化を推進する最初の力であったことを示唆しています。さらに、[[Rubicon]] フレームワークの前提と一致して、この機能は、サイズと複雑さが大幅に拡大したにもかかわらず、新皮質全体で適応学習を推進し続けます。このようにして、自然は哺乳類の脳における適応学習の 3 番目の柱を発見し、[[cerebellum]] と [[basal ganglia]] を超えて、よりオープンエンドで積極的に維持される [[bidirectional connectivity | bidirectionally connected]] システムへと移行しました。 |
一方、この中皮質リングの後半半分は、海馬と、空間ナビゲーションに不可欠な嗅内皮質や脾後皮質を含む関連ネットワークで構成されています ([[space]] および [[hippocampus]] を参照)。リングのこの部分の固定は、乳頭視床系との広範な相互接続によって部分的に生じ、その結果、長距離の空間ナビゲーションに重要な役割を果たすコンパスのような「頭方向」細胞が形成されます。さらに、このシステムは、この空間的およびエピソード的記憶ネットワーク内の活動を同期させる強力なシータ周波数振動 (10 hz、200 ms 周期) を駆動します。
この後部中皮質ネットワークには、後帯状皮質と、皮質系の単一の最も重要な空間表現領域である_後脾皮質_ (RS) が含まれます。これらは哺乳類の新皮質の「内側頭頂葉」ネットワークを構成しており、前方中皮質環との類推から、この空間ナビゲーション領域のネットワークは、あらゆる種類の空間運動処理に関与することが知られている背側(頭頂部)後新皮質の学習を固定している可能性があります。
典型的に夜行性のげっ歯類において、最も重要な感覚は嗅覚と体性感覚であり、これらの入力と空間的および目標駆動型の中皮質リングシステムが、げっ歯類の新皮質で起こることのほとんどを占めています。
霊長類や他の昼行性哺乳類では、視覚が支配的であり、ますます複雑化する視覚パターンと物体認識を処理するために必要な膨大な計算需要をサポートするために後頭葉が進化しています。したがって、一次視覚野 (V1) は新皮質の「基礎的な」領域であると考える人もいるかもしれませんが、実際には進化の歴史の中でははるかに最近のものです。その後、霊長類の側頭葉は、視覚物体認識と社会的コミュニケーションシステムを相互接続するシステムとしてさらに精巧になり、最終的にはヒトで発達した広範な言語能力をサポートします。
したがって、進化の軌跡を考察することは、他の脳領域と比較した新皮質の本質的な計算機能、およびこの領域の学習を形作る力を理解する重要な方法を提供します。
解剖学的詳細
{id=”図_構造” スタイル=”高さ:35em”}
![前頭前皮質領域の構造的構成。より多くの内側および腹側領域はより単純な無顆粒構造(つまり、進化的に古い中皮質)を持ち、より多くの背側領域と外側領域はより分化した複雑な真皮構造を持っています。 [[@^BarbasZikopoulos25]] より。](https://raw.githubusercontent.com/compcogneuro/web/main/content/media/fig_evolution_neocortex_structural_barbas25.png)
[[@^BarbasZikopoulos25]] は、中皮質リングの agranular (明確な第 4 層を欠く) 領域から、6 つの層すべてを持つ新皮質のより分化した eulaminate 領域への進行という観点から、[[prefrontal cortex]] 領域の structural 組織の包括的なレビューを提供します ([[#figure_structure]])。また、送信領域と受信領域の構造的特性の観点から理解できる皮質の相互接続性の違いも強調しています。
{id=”figure_ring2” style=”高さ:35em”}
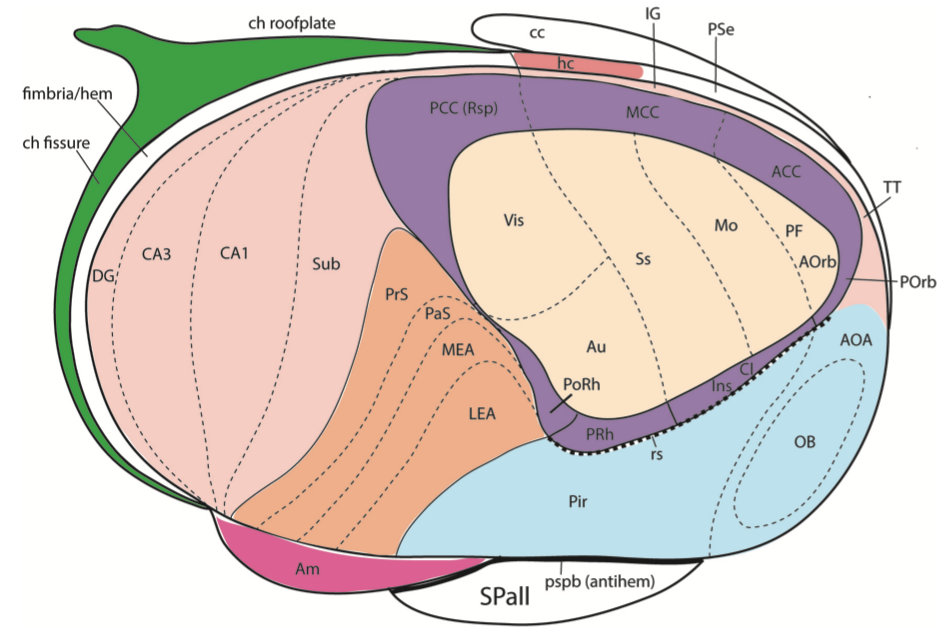
{id=”figure_embryo” style=”高さ:45em”}
![妊娠 9 週目のヒト胎児の中皮質環構造。[[@^AltmanBayer15]] より](https://raw.githubusercontent.com/compcogneuro/web/main/content/media/fig_evolution_neocortex_devgw9_ring_altmanbayer15.png)
[[#figure_ring2]] は、より精巧な二重リング構造 ([[@PuellesAlonsoGarcia-CaleroEtAl19]] より) を示しています。海馬とその関連サブフィールドが内側中皮質リング構造から区別され、内側中皮質リング構造には新皮質本来の異なる感覚コーディング領域が含まれており、完全な 6 層構造が出現します。 [[#figure_embryo]] は、妊娠 9 週目のヒト胚の中皮質環の構造を示しており、個体発生が系統発生を再現する (発生の軌跡は進化の軌跡を再現する) という原理を示しています。