compcogneuro/web: jang-et-al-2026
このページは外部資料の日本語訳です。原文の見出し順と本文順を保ち、コード・URL・出典表記はできるだけ原形のまま残しています。
出典とライセンス
原典: https://github.com/compcogneuro/web/blob/main/content/jang-et-al-2026.md
ライセンス: Text: CC BY 4.0; code: BSD 3-Clause。このページは日本語翻訳であり、変更点は翻訳とサイト内整形です。
+++ Name = “Jang et al (2026)” Title = “Synaptic Plasticity as a Function of the Temporal Derivative” Authors = “Jinyoung Jang1, Juan Flores1, Randall C. O’Reilly1,2, and Karen Zito1” Affiliations = “1Center for Neuroscience, University of California Davis, Davis, CA; 2Astera Institute” Abstract = “A major outstanding question in neuroscience is whether the neocortex somehow uses the same powerful learning algorithm that powers current AI models, which rely on error backpropagation. One hypothesized way this can be accomplished is as a function of the temporal derivative (i.e., differences in neural activity states over time), which can closely approximate the backpropagated error gradient. We tested this hypothesis that the direction of synaptic plasticity is a function of the temporal derivative in synaptic activity over the course of a 200 ms (5 Hz) theta cycle. Using a standard CA1 mouse slice preparation under patch clamp, we drove both pre- and postsynaptic activity across the two 100 ms halves of a 200 ms window at either 25 Hz or 50 Hz, testing all four 2x2 combinations of these firing rates, while measuring the resulting effects on synaptic efficacy (as measured by EPSP amplitude to standard test probes). Consistent with the computational hypothesis, a positive temporal derivative (25 to 50 Hz) resulted in LTP (increased synaptic strength), while a negative temporal derivative (50 to 25 Hz) resulted in LTD. Critically, both no-change conditions (25 to 25 and 50 to 50 Hz) resulted in no net synaptic change, even though the 50 Hz case had the highest overall synaptic activity levels. Possible biochemical mechanisms that could support these results are discussed.” Date = “2026-05-15” Categories = [“Papers”] bibfile = “ccnlab.json” +++ ## 導入
新皮質がどのように学習するかを理解することは、おそらく人間の知能を理解する上で最も重要なステップです。なぜなら、私たちの認知機能は、この脳構造内での長年の経験に基づく学習を経て現れるからです。この脳構造は哺乳類に特有であり、霊長類、特に人間で最も大きく拡張されています。現在の人工知能 (AI) システムは、強力な誤差逆伝播学習アルゴリズムに基づいています。この学習アルゴリズムは、人工ニューラル ネットワークにおける単一の最も有能な学習メカニズムとして長い間認識されてきました ([[@RumelhartHintonWilliams86]]、[[@WidrowHoff60]]、[[@Werbos74]])。したがって、このアルゴリズムは、新皮質がどのように学習すべきかについて最適な計算レベルの仮説を提供します。
誤差逆伝播を脳に実装する方法については、さまざまな提案が行われています ([[@LillicrapSantoroMarrisEtAl20]])。ここでは、そのような最も初期の提案の 1 つによる予測をテストします。この提案は、逆伝播誤差勾配が時間の経過に伴う神経活動状態の「時間導関数」によって近似できるという考えに基づいています ([[@OReilly96]]; [[@AckleyHintonSejnowski85]]; [[@XieSeung03]]; [[@ScellierBengio17]])。具体的には、活性化がボトムアップ方向とトップダウン方向の両方に流れることを可能にする双方向接続を備えたネットワークでは、ネットワーク内のニューロンのサブセットにわたるニューロン活動の変化がネットワークの残りの部分に反映されます。これらの変化が予測と正しい結果の違いを表し、時間微分に従って局所的なシナプス可塑性を駆動できる場合、結果として得られる学習は誤差逆伝播に近似します。
{id=”figure_bidir-err” style=”高さ:25em”}
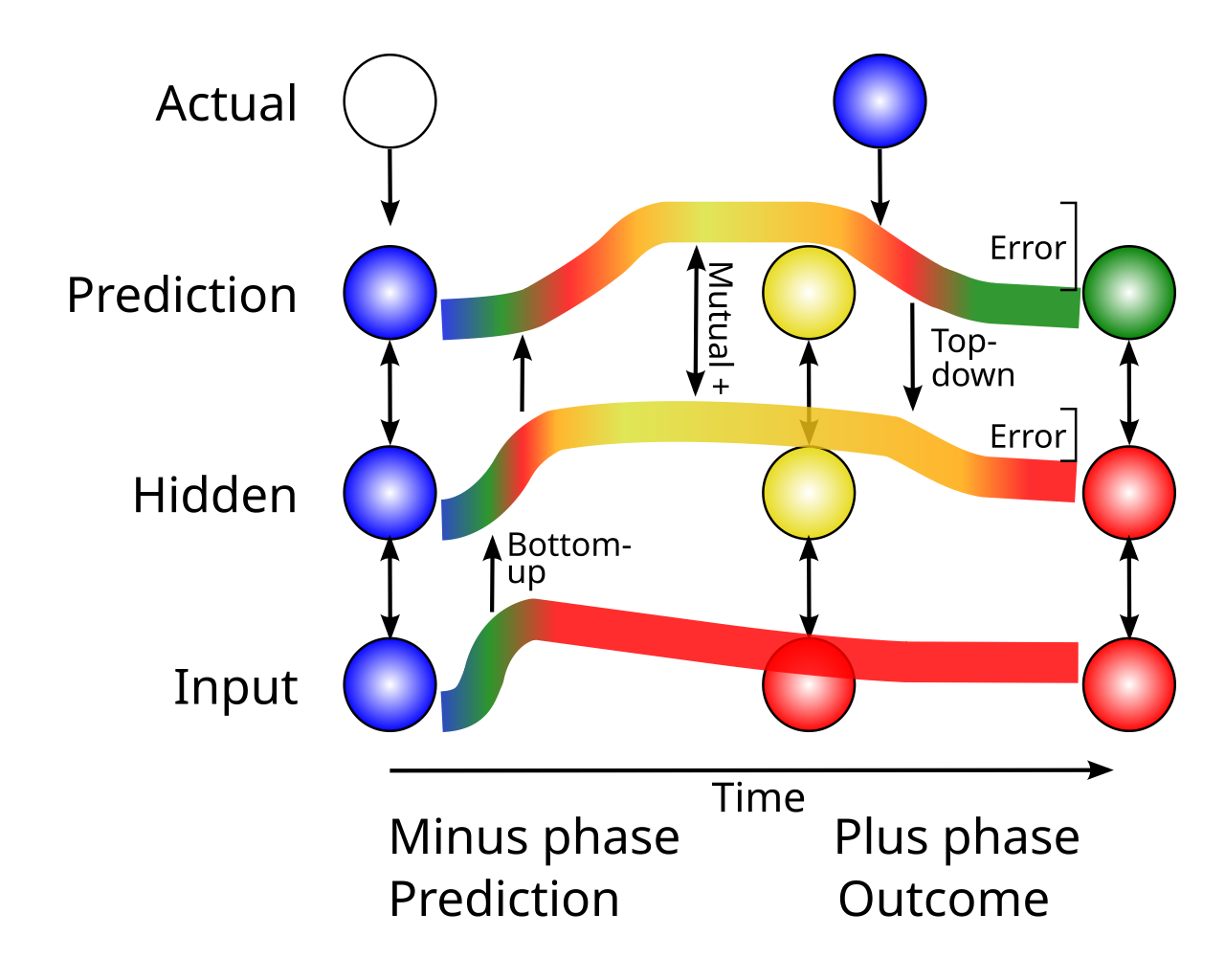
[[#figure_bidir-err]] は、これを単純な 3 層ネットワークのコンテキストで示しています。ニューラル活動の 2 つの異なる フェーズ は、シミュレートされたニューロンのような処理ユニットの入力層上に提示される特定の input パターンの影響を反映する初期の prediction フェーズまたは minus フェーズから始まります。続いて、「実際の」結果 (つまり、正しいまたは目標) アクティビティ パターンが予測レイヤーに駆動されると、アクティビティの「結果」フェーズまたは「プラス」フェーズが発生します。注目すべきことに、このようなネットワーク内の任意のニューロンでのこれらのアクティビティ状態 (plus_minus または outcome_predict、つまり、時間導関数 または 時間差) の単純な減算により、誤差逆伝播 ([[@OReilly96]]) によって計算される誤差勾配の良好な近似値が得られます。
したがって、このタイプの誤差駆動学習からの直接的な生物学的予測は、シナプス可塑性の方向が、予測段階と結果段階の間のこの移行を含む時間窓にわたる活動の変化(すなわち、時間微分)の関数であるはずであるということである。どちらも時間の経過に伴う変化に基づいているにもかかわらず、この新皮質学習メカニズムは、中脳内のドーパミン ニューロンの動作を記述する TD (時間差) 強化学習アルゴリズム ([[@SuttonBarto98]]; [[@MontagueDayanSejnowski96]]) とは完全に異なることに注意してください。 TD では、ドーパミン ニューロンは発火率の時間的な違いを「明示的に」表します。対照的に、新皮質の時間微分学習では、誤差勾配は時間の経過に伴う神経発火の変化に「暗黙的」に残りますが、この時間微分はあらゆる場所で局所的にシナプス可塑性を駆動します。誤差勾配のこの暗黙的な表現には、議論で詳しく説明したように、ニューラル計算を簡素化する上で重要な利点があります。
{id=”figure_pulv-conns” style=”高さ:15em”}
![一次視覚野と二次視覚野の場合、新皮質と視床の肺核の間の接続性は、予測エラー主導型学習を推進するのに非常に適しています。第 6 層 (VI) ニューロンからの多数の比較的弱い投射は、骨盤上の予測を活性化するのに適しており、複数の皮質領域およびニューロンからの信号を統合して予測を合成します。これは、学習の過程で新皮質全体で、およびこれらの最終投射で肺骨へ改善されます。対照的に、第 5 層(V)固有バースト(5IB)ニューロンからの強力な焦点ドライバー入力は、本質的に下位皮質層の活動パターンの未学習コピーである結果表現を活性化することができます(たとえば、この場合、V1 は V2 予測を訓練します)。 5IB ニューロンの周期的なバーストにより、この結果アクティビティは一時的にのみ存在し (つまり、プラス フェーズ)、完全な予測、つまり結果学習サイクルがおよそ 200 ms (つまり、シータ周波数、5 Hz) 以内に発生することが保証されます。 [[@^ShermanGuillery06]] に基づく図。](https://raw.githubusercontent.com/compcogneuro/web/main/content/media/fig_pulvinar_connectivity.png)
この学習アルゴリズムをさらに詳しく説明すると、この時間枠の期間に関する特定の仮説と、この形式の学習を駆動する予測および結果シグナルのソースに関する生物学的に明示的な仮説が提供されます ([[@OReillyRussinZolfagharEtAl21]])。具体的には、視床の新皮質と肺核の間にある視床皮質回路の独特な特徴が、200 ms (5 Hz) のシータ サイクルにわたって、予測→結果の状態 ([[#figure_pulv-conns]]) の交互シーケンスを駆動するはずです。この仮説は、[[@^OReillyRussinZolfagharEtAl21]] (例: [[@FiebelkornKastner21]]; [[@ShermanGuillery06]]; [[@ShermanUsrey24a]]) でレビューされているように、複数のレベルでの重要な証拠と一致しています。さらに、この同じシータサイクル時間微分学習メカニズムは、海馬の CA1 野における学習にも適用されます ([[@KetzMorkondaOReilly13]]; [[@ZhengLiuNishiyamaEtAl22]])。
{id=”figure_protocol” style=”高さ:30em”}
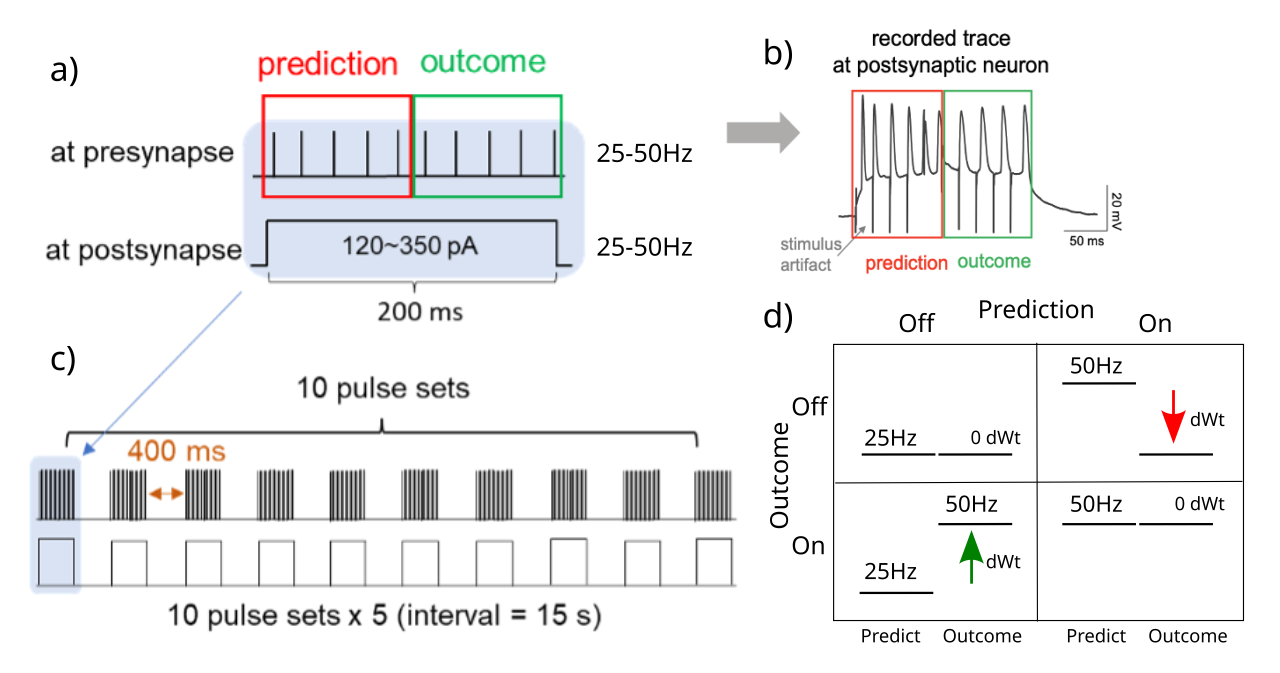
脳のシナプス可塑性が約200ミリ秒にわたる時間微分に敏感であるかどうかをテストするために、CA1領域と、除去されたCA3領域から始まるその求心性軸索線維を含むマウス脳スライスの標準的な実験準備を使用しました。個々の CA1 ニューロンが記録され、パッチ クランプ下で刺激されると同時に、軸索求心性神経も刺激され、これらの CA1 ニューロンへのシナプス入力における活動レベルの経時的な正確な実験制御が提供されました。我々は、200 ms シータ サイクル ([[#figure_protocol]]) の 2 つの 100 ms の半分にわたって、シナプス入力とクランプされたシナプス後 CA1 ニューロンの活動レベルを調整した方法で操作しました。
図に示すように、25 Hz と 50 Hz の低アクティビティ レベルと高アクティビティ レベルのすべての組み合わせを含む、予測 (前半) 対結果 (後半) アクティビティ レベルの 2x2 マトリックスがあります。時間微分アルゴリズムは、正の時間的変化 (つまり、結果 – 予測 > 0) は LTP (長期増強または正のデルタ重み (dWt) 変化) をもたらす一方、負の時間的変化は LTD (負の dWt) をもたらすはずであると予測します。さらに、シータサイクル全体にわたる活動の安定したパターンは、正味重量の変化がない (0 dWt) ことになるはずです。これは次の方程式に要約されます。
{id=”eq_dwt” title=”時間微分学習ルール”} \(dW = x^+ y^+ - x^- y^-\)
ここで、$x^+$ は結果 (プラス) フェーズでの送信ニューロンのアクティビティ、$y^-$ は予測 (マイナス) フェーズでの受信ニューロンのアクティビティなどです。この方程式は、複数の異なる開始仮定 ([[@AckleyHintonSejnowski85]]、[[@MovellanMcClelland93]]、[[@OReilly96]]) から導出でき、2 つのヘビアン $xy$ 因子の差またはコントラストであるため、「対照ヘビアン学習」 (CHL) 方程式と呼ばれています。
この学習ルールの対比的または時間微分的な側面は、一般にシナプス可塑性の方向と大きさが全体的なシナプス活動レベル $xy$ の関数であると予測する標準的なヘビアン学習メカニズムから定性的に区別するものです。歴史的に、シナプス前およびシナプス後活性の両方に敏感であるという NMDA 受容体の特性は、シナプス後末端ボタンの細胞内カルシウム レベルがこのヘビアン シナプス活性副産物を反映する、[[@^Hebb49]] ([[@DunwiddieLynch78]]; [[@Lisman89]]; [[@BearMalenka94]]) からの初期の理論的アイデアと一致することがすぐに認識されました。たとえば、全体的なシナプス活動の最大レベルは 50 Hz の安定したケースにあることに注意してください。この場合、時間微分メカニズムでは重み変化が 0 と予測されますが、標準的なヘビアン モデルでは正の LTP の最大レベルが予測されます。
以下の結果に示されているように、刺激プロトコルの下でのシナプス可塑性の方向は、時間微分学習ルールからの予測と完全に一致しており、したがって標準的なヘビアン学習メカニズムとは大きく矛盾していることがわかりました。以下では、シナプス変化の兆候に反対の影響を与える 2 つの化学経路間の比較的単純な競合結合力学がどのように時間微分学習特性を生み出すことができるかについて説明します。重要な特性は、増強経路が抑制経路と比較して全体的により速い応答時間定数を有し、自然に時間的導関数を生成することである。
## 材料と方法
パッチクランプ記録用の海馬スライスの準備
{id=”figure_slice” style=”高さ:25em”}
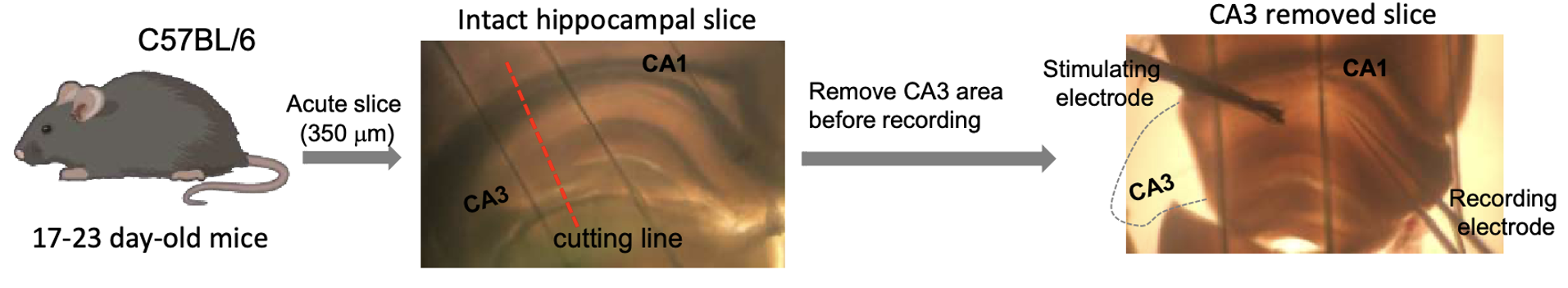
海馬スライスの記録には、生後 16 ~ 18 日目の C57BL/6 マウス ([[#figure_slice]]) を使用しました。マウスの脳を断頭後迅速に取り出し、氷冷した酸素添加人工脳脊髄液 (ACSF (mM): 127 NaCl、2.5 KCl、25 グルコース、25 NaHCO3、1.2 NaH2PO4、1 MgCl2、2 CaCl2、pH) 中で 320 $\mu m$ の厚さにスライスしました。 7.3) ビブラトーム (VT 1000S、Leica Microsystems) を使用します。回復期間中、急性マウス脳切片をACSF中で32℃で25〜30分間、室温で20〜30分間インキュベートした。
急性海馬スライスを作成するための詳細な手順:
-
実験当日、500 ml のパイレックスボトルに 250 ~ 300 ml の氷冷 ACSF を準備し、冷凍庫に約 20 ~ 30 分間、またはボトルの壁と表面に薄い氷の層が形成されるまで置きます。激しくかき混ぜて氷を砕き、均一な氷溶液にします。開始前に少なくとも 15 分間、95% O2/5% CO2 で氷冷 ACSF を氷中で酸素化します。
-
氷冷した ACSF を冷凍庫に入れている間に、スライス回収チャンバーに 100 ~ 150 ml の ACSF を準備します。開始前に少なくとも 15 分間、95% O2/5% CO2 で酸素を供給しながら、32°C の加熱バスでウォームアップします。
-
スライシングチャンバーを囲むトレイに氷を置き、ビブラトームを準備します。セラミックブレード (カタログ番号が見つかりません。Excel の注文シートに記載されています) をビブラトームのブレードホルダーに置きます。
-
ビブラトームの隣に解剖ツール (大きなハサミ、メス、細いハサミ、細い鉗子、片刃の刃、大きなペトリ皿 (100 mm)、ワットマン紙、接着剤、寒天ブロック、および湾曲したスパチュラ) をセットアップします。
-
酸素を添加した氷冷 ACSF の約半分 (150 ml) をペトリ皿に注ぎます。
-
大きなハサミを使用してマウスの首を素早く切り落とします。正中線に沿って皮膚を大きく切開して頭蓋骨を露出させ、両側の聴覚伝導を切断します。皮膚を動物の鼻に向かって引っ張って、頭蓋骨を完全に露出させます。
-
細いハサミで、小脳のすぐ尾側に切り込みを入れて頭蓋骨の後部を開き、尾側端から嗅球まで正中線に沿って頭蓋骨を切り開きます。頭蓋骨に圧力をかけないようにし、下のハサミの先端でその下の脳に損傷を与えないように注意してください。
-
細い鉗子を使用して、正中線の片側にある頭蓋骨の開いた端をつかみ、しっかりと保持して横に開き、もう一方の手で頭をしっかりと押さえます。次に、反対側に進みます。湾曲したスパチュラを使用し、非常に優しく脳の下に到達し、脳を優しくすくい出します。
-
抽出した脳をペトリ皿内の氷冷 ACSF に置きます。カッティングプレートに接着剤を適量塗布します。
-
片刃カミソリの刃を使用して、約 20 度の角度で脳の不要な部分を除去します。湾曲したスパチュラを使用して脳を拾います。スパチュラの底をペーパータオルの上にそっと置き、毛細管現象によって余分なACSFを排出します。スパチュラを接着剤の上に置き、脳を接着剤の上にそっと移します。スライスから脳をサポートするために寒天ブロックを配置します。
-
氷冷した ACSF をすぐにスライスチャンバーに注ぎ、95% O2/5% CO2 で酸素を供給し続けます。セラミックブレードが氷片に直接さらされるのを避けるため、メッシュを使用してスライスチャンバーの隅に氷を置いてください。
-
ビブラトーム コントロール パネルを使用してチャンバーをブレードに向かって持ち上げ、脳の表面に合わせてチャンバーの高さを調整します。 320 μm の海馬スライスを取得するためにスライスを開始します。
-
最初のスライスが解放されたら、両方の半球を分離し、加熱されたバス内の回収チャンバーに移します。すべてのスライスが収集されるまで繰り返します。
-
回収チャンバーを 32°C で 25 ~ 30 分間インキュベートします。回復チャンバーを加熱したバスから室温に慎重に移動し、記録を開始する前に 20 ~ 30 分間待ちます。スライスの回収後 4 ~ 5 時間は、実験にスライスを使用しないでください。
電気生理学
パッチ クランプ システム (Multiclamp 700B、Molecular Devices) を使用して、海馬 CA1 錐体ニューロンの膜電位を測定しました。記録電極 (3.5 ~ 4.5 M) は、Narishige プーラー (PC-10) を使用して、外径 1.5 mm のホウケイ酸ガラス キャピラリー (World Precision Instruments) から調製しました。 EPSP は、全細胞パッチクランプ構成の下、10 kHz のサンプリング レートで 1 kHz でフィルター処理された電流クランプ モードで記録されました。全細胞パッチクランプ記録の場合、KOHで調整したpH 7.3で、パッチピペットに135 K-グルコン酸塩、5 NaCl、10 HEPES、0.6 EGTA、4 Na-ATP、0.4 Na-GTP(mM単位)を充填しました。電気シナプス刺激には、ISO-Flex 刺激アイソレーターを備えた双極プラチナイリジウム微小電極 (FHC) を使用しました。
全細胞パッチクランプ記録の詳細な手順:
-
細胞内溶液を調製します。実験日を通して細胞内溶液を氷上に保ちます。
-
先端抵抗が 3.5 ~ 4.5 MΩ のガラス電極を引っ張ります。
-
アンプ (Multiclamp 700B、Molecular Devices)、デジタイザ (Digidata xxxx、Molecular Devices)、PC、ISO-Flex 刺激アイソレーター、デジタル カメラ システム、マイクロマニピュレーター (MP-285、MPC-200、WPI)、および顕微鏡コントローラーの電源を入れます。
-
酸素化 ACSF の流量を開始します。
-
広口の使い捨てピペットを使用して、海馬スライスの半分から領域 CA3 を除去した後、脳スライスを回復チャンバーから記録チャンバーに移します。
-
アンカー ワイヤーが海馬の CA1 領域に重ならないように、スライス ウェイトをスライスの上に置きます。
-
低倍率 (5 倍または 10 倍の対物レンズ) で CA1 錐体ニューロンを確認します。対物レンズを高倍率 (40 倍の対物レンズ) に交換し、錐体細胞の健康な領域を特定します。
-
低倍率レンズに戻り、双極刺激電極を配置します。刺激電極を領域 CA1 の放射状層に配置して EPSP を誘発し、刺激プロトコルを適用しました。刺激電極を配置するときは、電極の先端をターゲット CA1 錐体ニューロンから 5 mm 以上離して配置する必要があります。
-
全細胞パッチクランプが得られた後、正電流注入 (100 ~ 150 pA) を適用して活動電位を誘発します。刺激プロトコルに応じて、電流のサイズを調整して、異なる周波数の活動電位 (25 Hz または 50 Hz) を誘発します。
-
電流クランプ記録モードでは、電気刺激を加えて ~5mV EPSP を引き起こします。最も低い刺激強度から始めて、組織を焼いたり、刺激電極の先端で気泡を発生させたりすることなく、強度を上げていきます。最初の試行で ~5mV EPSP を誘発できない場合は、刺激電極をターゲット CA1 細胞に近づけてから、刺激強度のテストを繰り返します。各プローブ刺激の間には 15 ~ 30 秒の待ち時間を設けてください。
-
テストが完了したら、ベースライン EPSP の記録を開始します (5 ~ 6 分)。ステップ 9 と 10 に 5 分以上かかる場合は、このセルで実験を実行しないでください。別のセルを見つけて、手順 9 と 10 を繰り返します。3 ~ 4 個の障害がある場合は、スライスを変更します。
-
ベースライン EPSP 記録後、記録セルへの正電流注入と同じ電気刺激強度を使用して、誘導刺激パターンの 1 つを適用します。
-
EDL プロトコルが完了したら、すぐに EPSP の記録を開始します。 EPSP が記録されている間 (40 ~ 45 分)、負の電流注入 (-5 ~ 10 pA) によって EPSP 振幅と膜過分極を監視します。膜過分極の振幅が安定していない場合は、記録を停止し、スライスを変更します。導入プロトコルが適用される場合は、同じスライスを使用しないでください。
電気生理学データ分析
EPSP の振幅 (mV) は、各トレースの最初の 10 秒の膜電位 (mV) の平均であるベースライン膜電位からの膜脱分極のピークとして測定されました。これは、Clampfit 10.7 (Molecular Devices) で実行されます。アザラシと細胞の健康状態を監視するために、膜過分極もベースライン膜電位からの過分極の振幅として測定されました。
### 統計
統計分析は Excel (Microsoft office) を使用して実行されました。提示されたすべての数値およびグラフ表示は、平均値±平均値の標準誤差を表し、統計分析には両側 t 検定が使用されました。 P 値 < 0.05 は有意とみなされます。すべての統計はセル全体で計算されました。
刺激プロトコルの決定
{id=”figure_explore” style=”高さ:55em”}
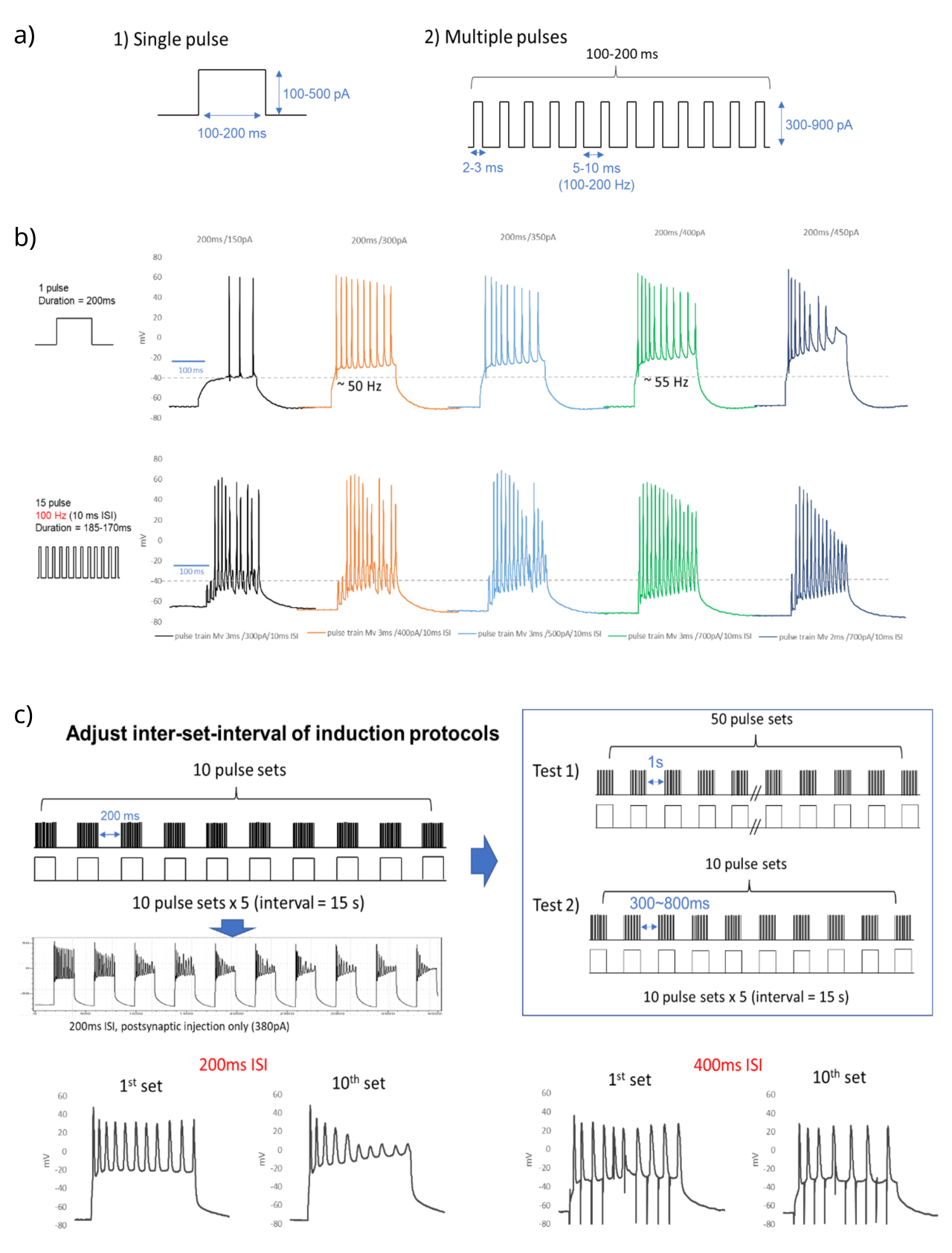
実験で使用される最終的な刺激パターンを決定するために、まず、一定の電流注入によって生成できる最大スパイク レートを決定しました。これは約 55 Hz ([[#figure_explore]]a および b) でした。また、個々の短い持続時間のパルス (3 ミリ秒) を使用して離散的な活動電位を駆動する可能性も調査しました。これにより、最大約 100 Hz の発火率が生成されます。安定した長時間 (200 ミリ秒) の電流注入を使用することにしました。これは、非常に多くの離散的な位相パルスの電流に比べてより自然であるはずであり、シナプス後ニューロンにおける 25 Hz と 50 Hz の低スパイク レートと高スパイク レートをサポートするには 55 Hz で十分であるためです。
次に、10 回の繰り返しにわたって明らかな劣化が生じなかった 200 ミリ秒のシータ ウィンドウ間のギャップを決定しました。 [[#figure_explore]]c に示すように、200 ms のギャップではスパイクの劣化が発生しましたが、400 ms のギャップでは劣化が起こらなかったため、400 ms のギャップを選択しました。 400 ミリ秒を超えるスパイク プロファイルに強い定性的変化は観察されませんでした。
## 結果
{id=”figure_results” style=”高さ:35em”}
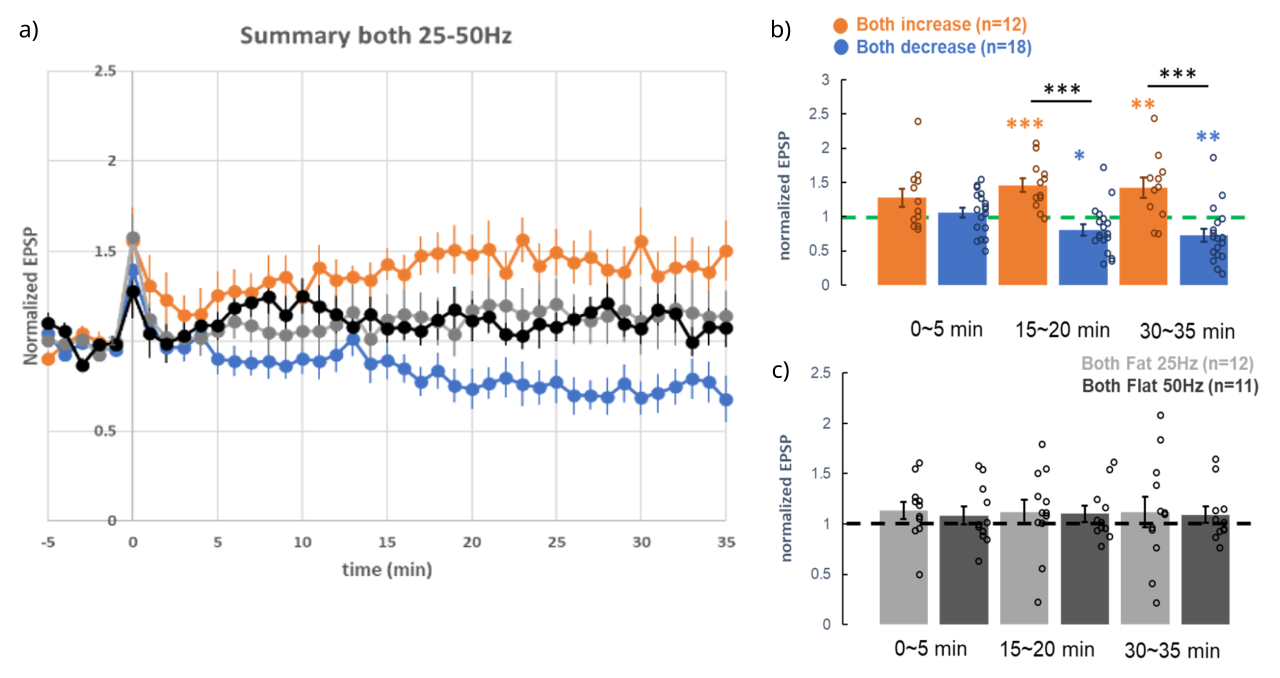
[[#figure_results]] は、[[#figure_protocol]] に示されている 4 つの時間微分条件すべての結果を示し、時間 0 で発生する誘導刺激プロトコル周囲の EPSP 振幅をプロットしています。シナプス前ニューロンとシナプス後ニューロンの両方が 25 Hz で 100 ms 駆動され、その後残りの 100 ms で 50 Hz に増加する条件では、LTP (30 ~ 35 分で正規化された EPSP) が発生しました。 = 1.42 ± 0.15、n=12、図中のオレンジ色の丸と棒)。逆に、時間微分値を減少させるプロトコル (50 ~ 25Hz) では、LTD (30 ~ 35 分での正規化 EPSP = 0.73 ± 0.10、n=18、図の青い丸と棒) が発生しました。最後に、両方のフラット プロトコル (一定の 25 Hz または 50 Hz) では EPSP 振幅に正味の変化はなく、25 Hz (30 ~ 35 分の正規化 EPSP = 1.12 ± 0.15、n=12、薄灰色の丸と棒) と 50 Hz (30 ~ 35 分の正規化 EPSP = 1.09 ±) の間に差はありませんでした。 0.08、n=11、濃い灰色の丸と棒)。
この結果のパターンは、時間微分学習メカニズム ([[#eq_dwt]]) の予測と完全に一致しており、一定の 50 Hz の場合が最も多くの LTP を示す一方、一定の 25 Hz が最も弱いか、LTD が生じるはずであると予測する既存のヘビアンのような学習ルールとは大きく「矛盾」しています。全体的に同じレベルのスパイクが、25 対 50 の場合と 50 対 25 の場合で 200 ミリ秒のシータ ウィンドウ全体に異なる分布をしているだけで、LTP と LTD (それぞれ) の反対のパターンが生成される可能性があるという考えは、標準的な Hebbian フレームワークの範囲を完全に超えています。
## 議論
私たちは、シナプス可塑性の方向は時間の経過に伴うシナプス活動の変化、つまり「時間導関数」の関数であるはずであるという仮説を検証しました。これは、強力な計算能力を持つ誤差逆伝播学習アルゴリズムの近似を実行できる能力と一致します。我々は、25 Hz と 50 Hz の 4 つの組み合わせすべてについて、2 つの連続する 100 ms ウィンドウにわたってシナプス前およびシナプス後活動の両方を駆動し、その結果生じるシナプス効率の変化をテストプローブで測定しました。 25 Hz から 50 Hz への活動の変化の増加は LTP (シナプス効率の増加) をもたらし、50 Hz から 25 Hz への減少の変化は LTD (シナプス効率の減少) をもたらすことを発見しました。一方、安定した 25 Hz または 50 Hz での両方の平坦な活動プロファイルでは、正味のシナプス効率の変化は生じませんでした。
このシナプス効率変化のパターンは、GeneRec ([[@OReilly96]]; [[@XieSeung03]]; [[@ScellierBengio17]] も参照) として知られる誤差逆伝播に対する時間微分ベースの近似の予測と完全に一致しています。これは、Recirculation アルゴリズム ([[@HintonMcClelland88]]) の一般化であり、位相ベースの学習に基づいています。このアイデアは最初に Boltzmann Machine ([[@AckleyHintonSejnowski85]]) で開発されました。したがって、ここで報告された結果は、新皮質がこの脳領域に独自に存在する確立された双方向の興奮性接続を利用して、誤差逆伝播を介して学習するという仮説に対する重要な経験的裏付けを提供します ([[@VanEssenMaunsell83]]; [[@MarkovErcsey-RavaszLamyEtAl13]])。この双方向接続により、新皮質のどの部分でも活動の変化が広範囲に伝播し、それによってエラーの逆伝播と同じ効果が達成されます。
さらに、新皮質と高次視床核(髄核および中背核)との間の視床皮質接続の独特の特性は、予測と結果の活動状態の交互フェーズ([[@OReillyRussinZolfagharEtAl21]])を確実に駆動するためのメカニズムを提供します。これは、時間微分ベースの誤差逆伝播学習にとって必須の要件です。したがって、複数のレベルの分析で利用可能な神経生物学的データは、この強力な計算能力を備えた学習形式の要件と全体的に一致しています。
ヘビアン学習との比較
文献における新皮質学習の主な計算レベルの解釈は一般に、NMDA受容体を介して入るシナプス後カルシウムのレベルとシナプス可塑性の方向と大きさとの関係を示す十分に確立されたデータに基づいて、さまざまな形のヘビアン学習に焦点を当ててきた([[@Lisman89]]; [[@BearMalenka94]])。具体的には、カルシウムのレベルが低いとLTDが発生し、カルシウムのレベルが高いとLTPが発生します。これは一般に、ヘビアン学習アルゴリズムの BCM ([[@BienenstockCooperMunro82]]) バージョンと一致しています。
最近では、スパイクタイミング依存可塑性 (STDP) ([[@BiPoo98]]) が計算モデルの主な焦点となっています (例: [[@KheradpishehGanjtabeshThorpeEtAl18]]、[[@DiehlCook15]])。しかし、最初に説明された STDP の単純な計算強制形式は、1 秒間隔で分離された個々のスパイクのペアによる非常に特殊な刺激プロトコルを必要とし、一般に、より現実的な神経活動パターンには適用できないことが明らかになりました ([[@DebanneInglebert23]])。実際、同じ BCM のようなパターンが、より現実的で密度の高いアクティビティ パターンで現れます ([[@ShouvalWangWittenberg10]])。
これらのヘビアン学習メカニズムに対する誤差逆伝播の計算上の重要な利点は、ネットワーク内のすべてのニューロンにわたる学習を調整し、遠位誤差信号を最小限に抑えるように数学的に設計されていることです。対照的に、Hebbian 学習には、共活性化の統計的規則性を抽出するという点で局所的なヒューリスティックな機能しかありません ([[@Oja82]]; [[@RumelhartZipser85]]; [[@IntratorCooper92]])。したがって、ヘビアン学習が新皮質に存在するような深層ネットワークを効果的に訓練できると信じる理由はありませんが、これはまさに誤差逆伝播が優れている場合です。したがって、今回の結果は、新皮質学習の計算レベルの要求とその根底にある神経機構とを調和させる上で重要な潜在的な方法を提供する。
さらに、今回の結果は、LTP と LTD に関する実験文献の比較的理解しにくいいくつかの特徴についての説明を提供する可能性があります。特に、実験的にLTPを生成するために必要なシナプス活動のレベルは生理学的に過剰になる傾向があり、一方でLTDを確実に得るのは一般に比較的困難でした([[@BearAbraham96]])。これらの既存の刺激パラダイムは、シナプス可塑性の方向を駆動するために重要であることがわかった 200 ms ウィンドウにわたる種類の時間的変化を体系的に誘発していません。したがって、おそらく、この論文で調査した種類の時間ダイナミクスをさらに調査することで、より実験的に信頼できる形態のシナプス可塑性が得られる可能性があり、これは純粋に実用的な理由からも有益となるでしょう。
競合するキナーゼを介した時間的誘導可塑性
私たちが得た結果は、シナプスで作用する既知の基礎的な生化学プロセスからどのようにして得られるのでしょうか?数学的には、時間微分は、共通の駆動入力信号の fast 積分から slow 積分を引いたものの差として計算できます。直観的には、高速積分はより最近の結果状態をより厳密に反映しますが、低速積分は以前の予測状態からの痕跡をより多く保持します。この原則の対話型デモンストレーションについては、compcogneuro.org の [[temporal derivative]] を参照してください。
神経化学的に、LTP と LTD の違いは、2 つの異なる_キナーゼ_、CaMKII (カルシウム カルモジュリン キナーゼ II) と DAPK1 (死関連プロテイン キナーゼ 1) 間の競合によって部分的に決定されます。これらのキナーゼは両方ともカルシウム活性化カルモジュリン (CaM) (([[@GoodellZaegelCoultrapEtAl17]];[[@GoodellZaegelCoultrapEtAl17]]; [[@GoodellTullisBayer21]]; [[@TullisBayer23]]; [[@BayerGiese25]])、CaMKII の方が一般的な CaM ドライバーの全体的な統合が高速であり、DAPK1 の統合がより低速である場合、これは必要な時間微分メカニズムを実装します。
したがって、全体的な統合率のこの違いは、この全体的な枠組みからの予測として存在しており、より一般的には、ここで報告するシナプス可塑性の形態の根底にある神経化学プロセスと、まだ調査されていない神経活動信号の考えられるさまざまな違いに対するその感度について、多くの未解決の主要な疑問が残っています。
皮質ダイナミクスと予測コーディング
皮質視床予測対結果メカニズム ([[#figure_pulv-conns]]; [[@OReillyRussinZolfagharEtAl21]]) によってサポートされるエラー駆動学習は、広く議論されているベイジアン予測コーディング フレームワーク (例: [[@RaoBallard99]]; [[@Friston09]]) の代替手段となります。この代替案の時間微分基礎により、予測学習をサポートするために必要な皮質ダイナミクスが大幅に簡素化され、利用可能なデータとよりよく一致します。
ベイジアン モデルでは、ボトムアップの実際の結果からトップダウンの予測を減算することによって、ニューロンの部分母集団が_予測誤差_を明示的に表す必要があります。したがって、異なるニューロン集団が基本的に異なる情報を表現できるように、何らかの方法で分離する必要があります。さらに、これら 3 つの異なる信号 (予測、結果、エラー) はすべて、原理的には層を越えて異なる方向に伝達される必要があり、強力に分離された経路が必要です。
対照的に、時間微分モデルでは、ネットワーク全体は、どの時点でも常に「一貫性」があり、「相乗的」です。すべての層とニューロンが基本的に協力して、世界の「現在の」状態の一貫した解釈を表します。この現在の状態は、時間の経過とともに、予測と結果の表現が交互に繰り返されるだけです。結果が予測と一致する場合、変化はありません。これは通常、成熟し、よく訓練されたシステム、つまり安定した正確な世界の表現における状況です。ただし、発達学習の初期や、成熟したシステムにおける比較的新しい状況や困難な状況では、予期せぬ結果によって学習が促進され、予測状態の精度が向上することがあります。
したがって、この形式の学習により、ネットワーク内のすべてのレベルが連携して並列の制約満足処理を推進し、トップダウン制約とボトムアップ制約を統合して、現在の状態の一貫した解釈を推進することができます ([[@HopfieldTank85]]; [[@OReillyWyatteHerdEtAl13]])。これは、表現空間を介した [[search]] の強力な形式を表し、システムの予測精度を向上させるために、シナプス重み空間を介した誤差逆伝播探索の外側ループ内の一種の内側ループ最適化として動作します。 [[@^OReillyRussinZolfagharEtAl21]] および compcogneuro.org で広く報告されている計算モデルは、生物学的に現実的なスパイキング ニューロンを使用した、この形式の学習と処理の有効性を実証しています。
入手可能な神経学的証拠は、皮質のすべてのレベルにわたる情報の一貫性、相乗的、冗長エンコーディングと一致しており、ベイジアン モデルで必要とされる種類の構造的分離に関する重要な証拠はありません ([[@WalshMcGovernClarkEtAl20]]; [[@HeilbronChait18]])。発見された主要な肯定的な証拠、つまり予期せぬ結果に対する期待される結果に対する神経活動の抑制は、十分に確立された神経適応/順応メカニズムと組み合わせた代替時間微分モデルと互換性があります ([[@KokLange15]]; 詳細な議論については [[@OReillyRussinZolfagharEtAl21]] を参照)。
したがって、時間微分フレームワークは、新皮質が、複数のレベルの分析で利用可能な神経証拠とより互換性があると思われる方法で、次に何が起こるかについてトップダウンの予測を生成することによって学習するという広く受け入れられている考えをサポートしています。
### 結論
結論として、ここで提示された結果は、新皮質学習が、時間導関数によって駆動されるシナプス可塑性に基づいて、計算的に強力な誤差逆伝播学習メカニズムを実装している可能性をテストするための重要な第一歩を示しています。この形式の学習は、既存の神経科学データの広範なデータと一致しており、神経科学における最も重要な未解決の疑問の 1 つに対する考えられる答えをより直接的に調査するためのさらなる実験的研究から恩恵を受けるでしょう。