compcogneuro/web: motor
このページは外部資料の日本語訳です。原文の見出し順と本文順を保ち、コード・URL・出典表記はできるだけ原形のまま残しています。
出典とライセンス
原典: https://github.com/compcogneuro/web/blob/main/content/motor.md
ライセンス: Text: CC BY 4.0; code: BSD 3-Clause。このページは日本語翻訳であり、変更点は翻訳とサイト内整形です。
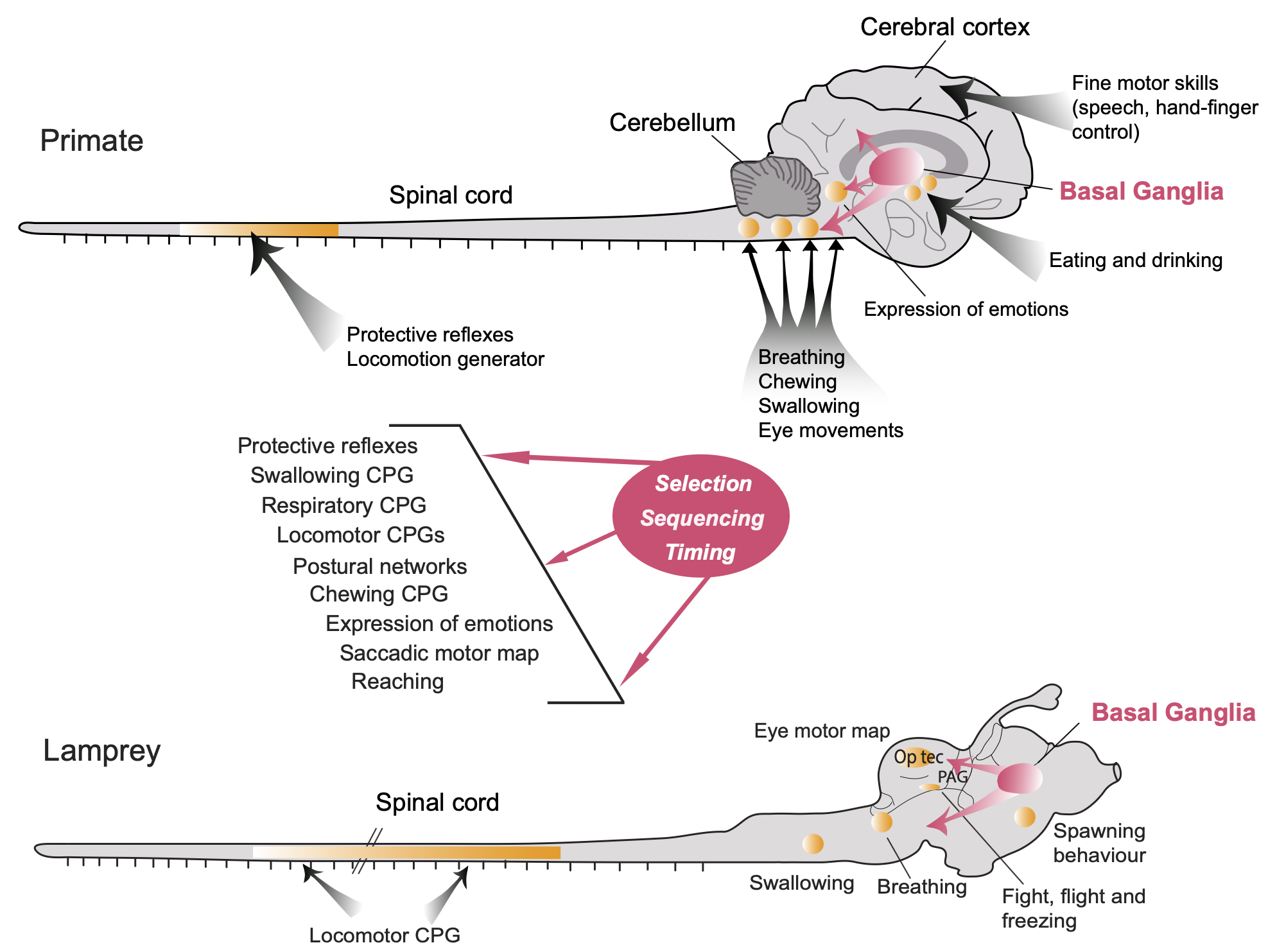
+++ Categories = [“Neuroscience”, “Cognition”] bibfile = “ccnlab.json” +++ 脳の 運動 システムには多くの異なる領域が関与しており、さまざまな方法で脳全体が運動出力を生成するために存在しています ([[#figure_overview]])。たとえば、ダニエル・ウォルパートは、ホヤは動き回る必要がなくなったら自分の脳を食べると主張しました。モーター制御のさまざまな側面のより具体的な説明は、[[reinforcement learning]]、[[basal ganglia]]、[[cerebellum]]、[[space]]、および [[Rubicon]] フレームワークにあります。このページでは、運動制御の課題と解決策の高レベルの概要を提供し、筋肉の基礎的な生理機能と、脊髄と脳幹が運動制御の体系的な基盤をどのように提供するかについての詳細を示します。
{id=”figure_overview” style=”高さ:35em”}
次元の削減、調整、筋肉の相乗効果
神経機能のあらゆる側面と同様に、運動制御における中心的な問題は [[curse of dimensionality]] を管理することです。時間の経過とともに、筋肉の活性化には指数関数的に多くの可能な組み合わせが存在します。脳はどのようにしてこの巨大な空間を、動物の生存に必要な行動を実行するために必要な筋肉活動の比較的小さなサブセットにまで縮小するのでしょうか?
これは基本的な [[search]] 問題のもう 1 つの例であり、運動システムの場合、[[evolution]] は脊髄と脳幹に複雑な 筋肉の相乗効果を組み込むことで多くの仕事を行い、介在ニューロン (興奮性および抑制性) のネットワークが筋肉活性化の 時空間パターンを活性化します。この低次元の要素 (つまり [[linear algebra#basis space]]) から、運動動作の複雑なパターンが構築されます ([[@Bernstein67]]; [[@Bernstein96]]; [[@TreschSaltielBizzi99]]; [[@dAvellaSaltielBizzi03]]; [[@TingMcKay07]]; [[@BizziCheungdAvellaEtAl08]]; [[@TreschJarc09]]; [[@OverduindAvellaRohEtAl15]]; [[@Latash20]];
これらの筋肉の相乗効果によって解決されるもう 1 つの重要な問題は、特定の運動動作を達成するために必要な、多くの異なる筋肉にわたる 調整 です。特定の筋肉の収縮は、生物全体の基本的な重心の変化など、他の筋肉にさまざまな物理的影響を引き起こします。したがって、特定のアクションでは、これらの結果をすべて考慮し、個々の筋肉の収縮がすべて実際に相乗的であり、交差した目的で機能しないことを確認する必要があります。
文献では、これらの相乗効果を指すために、反射 ([[@Sherrington10]])、中枢パターン生成装置 (CPG、[[@GrillnerElManira20]]; [[@GrillnerZangger79]])、および_力場_ ([[@GiszterMussa-IvaldiBizzi93]]) など、さまざまな異なる用語が使用されています。特に CPG の場合、これらの用語には多少異なる意味合いがあり、これは、たとえば長期間にわたる持続的でリズミカルなパターン化された行動を含む、運動行動を駆動する複雑な回路に当てはまります。より一般的には、これらの相乗効果は、モジュール ([[@BizziCheungdAvellaEtAl08]])、プリミティブ、ビルディング ブロック、または運動動作のコンポーネントと呼ばれます。
{id=”figure_synergies” style=”高さ:30em”}
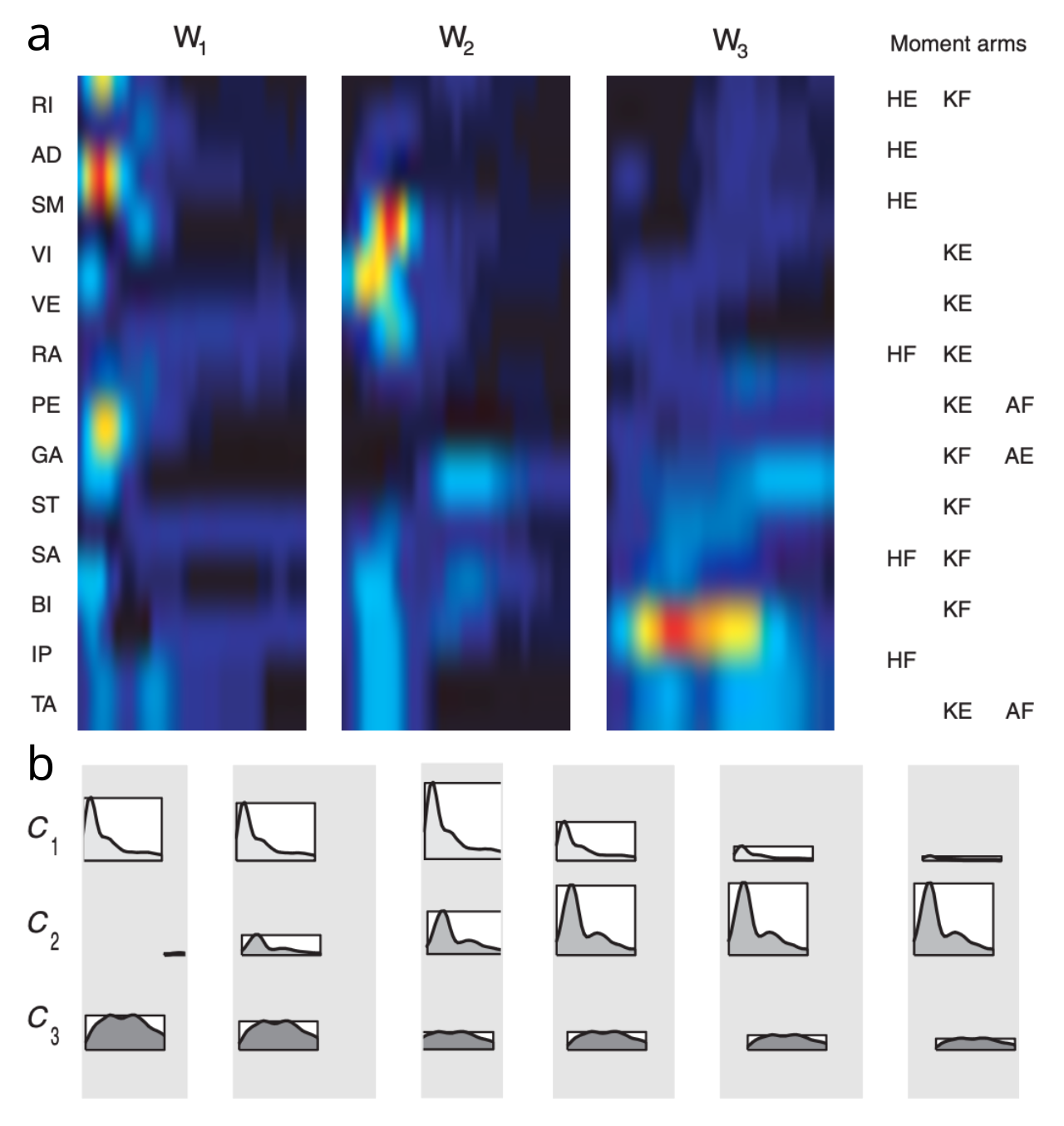
具体的な例として、カエルの脚系における脊髄レベルの筋肉の相乗効果の詳細な分析により、さまざまな活性化強度と組み合わせて広範囲の全体的な運動反応パターンを説明できる 4 つの相乗効果のセットが明らかになりました ([[@TreschSaltielBizzi99]])。無傷のカエルの脳全体からの寄与を考慮したその後の分析では、3 つの異なる時空間筋肉の相乗効果 ([[#figure_synergies]]) のタイミングと活性化の調節が、この同じ運動反応空間 ([[@dAvellaSaltielBizzi03]]) をどのように説明できるかが示されました。
したがって、最終的な運動行動パターンは、複数の制御レベルの統合された貢献に依存しており、介在ニューロンによって実行される脊髄筋の相乗効果が最低レベルを提供し、より高いレベルでは、独自の直接的な相乗効果と、基本的な脊椎要素全体にわたるより広範な調整が徐々に追加されます([[@GrillnerElManira20]])。たとえば、手の動きにおける相関する運動単位の分析では、基本的な脊髄介在ニューロン ([[@XuMawaseSchieber24]]) に加えて、橋髄網様体、大細胞性赤核、一次運動野 (M1) からの_最終順序_ (つまり、筋肉活動のすぐ上流) の入力が示されました。
これらの筋肉の相乗効果の遺伝的にコード化された性質は、研究対象となった個々の動物のそれぞれに同じ相乗効果が見出され(例: [[@dAvellaSaltielBizzi03]]; [[@BizziCheungdAvellaEtAl08]])、人間においても同様の相乗効果が見出され、成人期まで持続する([[@YangLoganGiszter19]])ことを示す多くの研究で明らかであり、移動の中心的なパターンは他の種のものと強い類似性を持っています。 ([[@DominiciIvanenkoCappelliniEtAl11]])。また、汎用学習メカニズムは通常、筋肉の相乗効果に似た低次元の基底空間を開発しますが ([[@ChaiHayashibe20]]; [[@TreschJarc09]]; [[@Scott04]]; [[@TodorovJordan02]])、脊椎実装の直接的な証拠と強力な遺伝的証拠は、動物におけるこれらの進化的基盤を裏付けています。
感覚フィードバック制御
運動システムにおける次元削減に対するもう 1 つの重要なニーズは、動物とその環境の間の相互作用から生じます。これも非常に変化しやすいため、分散と組み合わせ爆発の主要なさらなる原因となります。脊髄介在ニューロンによって実行される筋肉の相乗効果には、環境の摂動を自動的に補償する直接フィードバック制御メカニズムが組み込まれているというかなりの証拠があります ([[@WimalasenaPandarinathAuYong25]]; [[@ShinoharaAmbeKimEtAl25]]; [[@ConwayHultbornKiehn87]]; [[@AngelGuertinJimenezEtAl96]]; [[@AlvarezFyffe07]]; [[@BizziCheungdAvellaEtAl08]]; [[@SantuzAkayMayerEtAl19]]; [[@GorkoSiwanowiczCloseEtAl24]])。ある推定では、これらのフィードバック信号の力の寄与は 35% とされています ([[@SteinMisiaszekPearson00]]; [[@CoteMurrayKnikou18]])。
残念ながら、これらのフィードバック制御メカニズムを理解する能力は、これらのフィードバック信号を処理する脊髄介在ニューロンの接続性を分類およびマッピングすることが困難であるために損なわれています ([[@SenguptaBagnall23]]; [[@McCrea92]]; [[@CoteMurrayKnikou18]])。さらに、現存する回路ベースのモデルにそのようなメカニズムが組み込まれ始めたのは比較的最近になってからです ([[@ShinoharaAmbeKimEtAl25]] – [[#figure_spinal-cpg]] および [[@DannerShevtsovaFrigonEtAl17]] を参照)。それにもかかわらず、大規模な脊髄介在ニューロン集団のより抽象的な状態空間解析は、これらのフィードバック制御メカニズムの存在と力学的意味を明らかにする可能性を秘めています ([[@WimalasenaPandarinathAuYong25]])。
運動システムにおけるフィードバック制御の理解はかなり未発達ですが、知覚制御理論 (PCT) ([[@Powers73]]; [[@Powers73a]]; [[@Cools85]]; [[@Yin14a]]; [[@BarterYin21]]; [[@ParkerWillettTysonEtAl20]])。このフレームワークの中心的な前提は、制御が感覚信号に基づいて動作する必要があるということです。なぜなら、これらの感覚信号はシステムの_関連する_状態を知らせてくれるからです。
簡単な例として、冷暖房システム (HVAC) は、必要な熱または冷却の量は明らかに「環境」に依存するため、目標の熱量または冷却出力 (「モーター」変数) ではなく、目標の温度 (感覚変数) によって駆動される必要があります。たとえば、古い車は HVAC 出力の制御のみを提供し、ユーザーが必要な感覚変数との接続を行うことに依存していましたが、最近の車では目標温度によって駆動されるサーモスタット制御が搭載されており、制御タスクが大幅に簡素化されています。
つまり、特定の行動の結果を達成するために使用できる環境パラメータと運動パラメータのさまざまな組み合わせが存在するため、これらの運動パラメータのレベルで制御を行うことは、望ましい結果に「直接」関係する感覚変数を制御することに比べて、十分に決定されておらず、非効率的です。
PCT の経験的実証は、運動システムが混乱しているときに人々が行っている実際の運動動作についての認識が完全に欠如していることを示しています ([[@ParkerWillettTysonEtAl20]])。いずれにせよ、脊髄レベルでの運動相乗効果の存在を考慮すると、運動制御が個々の筋肉または関節ベースの信号から分離されているのはすでに事実です(ただし、以下で検討するように、霊長類、特に人間の運動皮質には、よりきめの細かい筋肉レベルの制御を提供する能力があります)。
モータ制御に密接に関連し、影響力のある理論は 最適フィードバック制御 (OFC) です。これも同様に、環境やその他の変動要因に直面した場合、オンラインの感覚フィードバック ベースの制御が唯一妥当な制御形式であるという原則に基づいています ([[@TodorovJordan02]]; [[@Scott04]])。 PCT と同様に、OFC は、力などのモーター変数を直接駆動するのではなく、制御がフィードバック ゲイン パラメーターを調整するという原理に基づいています。
OFC は、生物学的な制約によって実際のシステムがさまざまな点で少なくともある程度最適化されていない可能性があるにもかかわらず、有用な参照点として、計算レベルの最適化メカニズムとの重要な関係を追加します。これらの最適化メカニズムの結果の 1 つは、目標に関連する結果 (たとえば、上の例の温度) のみに焦点を当て、他の無関係な結果は無視することです。これは経験的データ ([[@VazPintoJuniorEtAl19]]) と一致しています。
非制御多様体 (UCM) フレームワークは、最適化メカニズムの関与を識別するための、紛らわしい名前ではありますが、広く使用されている手法を提供します ([[@ScholzSchoner99]]、[[@LatashScholzSchoner07]]、[[@Latash24]])。直感的には、最適化プロセスによって制御されている変数 (温度など) は、controlled パラメーター (熱と冷却の出力レベルなど) のようなラベルが付けられる一連の基礎となるパラメーターに関連付けられると予想されます。ただし、これらは実際には、uncontrolled 多様体内の変数です。なぜ?なぜなら、通常は、同じ望ましい制御された出力を達成するために、さまざまな「非制御」方法で設定できる複数の異なるパラメーターが存在するからです。
たとえば、2 つの異なる熱源 (変化が遅い受動的床暖房と、変化の速い強制換気口など) がある場合、これら 2 つの熱源のさまざまな組み合わせがすべて UCM を表します (ここで、多様体 とは、考えられるすべてのパラメーター値のより大きな空間内の制約された部分空間を指します)。任意の時点で、これらの熱源の特定の組み合わせによって目標温度を達成できます (たとえば、電流を考慮し、床熱をゆっくりと変化させ、必要に応じて強制換気口を調整します)。一方、冷却源 (およびその他の考えられる無関係なパラメーター) はすべて、科学実験における制御変数という意味で「制御」されています。つまり、操作されている関連する「独立」変数に干渉しないように分散が最小限に抑えられているものです (この場合、「独立」は実質的に「制御されていない」の同義語です)。
紛らわしい名前にもかかわらず、UCM フレームワークは、モーター制御における「冗長な」モーター パラメーターの完全な部分空間多様体を持つことが、モーター制御における重要な種類の「豊富さ」 ([[@GelfandLatash98]]) を表すという重要なアイデアも捉えています。そこでは、これらのさまざまなパラメーターを実際に活用して (あえて「制御される」と言うかもしれません)、斬新で複雑な状況 (例: 異なる手の構成で同時に複数のものを運ぶなど) に適応するための重要な 柔軟性 を提供できます。物を運ぶときのドアなど)。したがって、これらすべての新しい状況にその場で適応できる、十分に強力で柔軟かつ動的なオンライン コントローラー メカニズムがあれば、次元の呪いを問題とみなすのではなく、チャンスでもあります。これらの方針に沿った UCM とマッスル シナジー フレームワークとの統合については、[[@^LatashScholzSchoner07]] を参照してください。
姿勢と平衡点の制御
PCT フレームワークは、平衡点 運動制御の原理と一致しています。この原理では、力などの動的変数ではなく、下降する運動制御信号が各筋肉の_目標長_を指定すると仮定しています ([[@Feldman86]]; [[@FeldmanLevin09]]; [[@GribbleOstrySanguinetiEtAl98]]; [[@Sainburg15]])。
平衡点モデルの魅力的な利点の 1 つは、姿勢 (座る、立つなど) と能動的な運動動作との関係を理解するための統一されたフレームワークを提供することです。この文脈における姿勢は、筋肉の長さと関節角度の固定された構成にすぎず、動作はこれらの姿勢パラメータの更新にすぎません。目標の長さが変更されると、自動フィードバック筋力が作用して、システムを新しい目標の長さに移行させます。したがって、このフレームワークでは、_アクション_は単に姿勢を変更するだけです。
特定の姿勢を維持するには、継続的な筋肉の活性化が必要です。平衡点モデルによれば、下降制御システムが対応する能動制御入力を維持する必要があるのではなく、そのようなシステムが望ましい姿勢構成を確立し、関連するフィードバック機構によって駆動される脊髄レベルの回路がこの目標構成を維持できるようにする方が合理的である。
しかし、脊髄に存在する筋肉の相乗効果が実際に感覚フィードバック機構 (筋紡錘線維によって信号が送られる筋肉の長さを含む) を組み込んでいることが理解されると、このフレームワークと平衡点モデルの間の境界は溶解し始めます。それにもかかわらず、筋肉の相乗効果に対するこれらの感覚フィードバック信号の重要性は、下降制御を簡素化し、筋肉の相乗効果の堅牢性と適応性を向上させるために重要であることを考えると、本来あるべきほど広く認識されていない可能性があります。
平衡点フレームワークの精神と一致して、脊髄には固有受容フィードバック信号に反応する複数の異なる姿勢制御「反射」(相乗効果)の証拠があります([[@TingMcKay07]]; [[@DeliaginaZeleninBeloozerovaEtAl07]]; [[@EliasWatanabeKohn14]]; [[@BinghamChoiTing11]])。たとえば、[[@^BrambillaRussodAvellaEtAl23]] は、緊張と活動の姿勢相乗効果の小さなセットの証拠を発見しました。しかし、これらの姿勢の相乗効果は、さまざまな位相的動作に関与する相乗効果とは大きく異なり、すべての動作は単なる姿勢の変化であるという考えに反しています。
平衡点フレームワークのもう 1 つの中心点は、特定の姿勢を維持することと、特定の運動動作を活性化すること (姿勢を乱すことになる) の間に潜在的な矛盾があることです。筋肉相乗効果の枠組みでは、これは、特定の時点で活性化する可能性のあるすべての筋肉相乗効果要素の中から選択するという、より広範な課題の一例として見ることができます。これらの低レベルの相乗効果に感覚フィードバックが直接関与することで、同時にアクティブな他の相乗効果の存在に適応することができますが、これらの相乗効果をより適応的に調整するには、次に説明するように、より高いレベルの制御が必要になる可能性があります。
制御レベル
最下位レベルの脊椎相乗作用は、脳幹、小脳、大脳基底核、および新皮質の運動核から下降制御入力を受け取ります ([[#figure_overview]])。これらのより高いレベルの制御の具体的な貢献をどのように理解できるでしょうか?
まず、純粋に空間的な相乗効果 (つまり、筋肉が活性化される静的なパターン) と、時間的な要素も含まれる相乗効果との間には重要な違いがあります。この区別は、論理的には、相乗効果自体のレベル (つまり、特定の相乗効果が生成する筋肉活性化の性質) と、全体的な動作を生み出すためにこれらの相乗効果を駆動する入力制御信号のレベルで行うことができます。 [[#figure_synergies]] に示されている例では、モデルはこのように制約されていませんが (したがって、両方に時間コンポーネントが存在します)、制御信号は本質的に静的である可能性があるように見えますが、時間的なダイナミクスはすべて相乗効果自体内に吸収されます。
脊椎シナジーネットワークにおける固有受容および他の体性感覚入力からの局所感覚フィードバック制御の中心的な役割は、これらのシナジー内で生じる時間的ダイナミクスの自然な基盤を提供し、それによって制御信号をより単純な強直性活性化入力とすることができる(例えば、平衡点フレームワークの精神において)。さらに、移動やその他のリズミカルな行動に関連する CPG のような相乗効果の場合、明らかに脊髄プリミティブが重要な時間成分に寄与する一方で、制御信号は移動の全体的な速度を調節することが示されています ([[@Grillner81]]; [[@GrillnerElManira20]])。
しかし、脊椎プリミティブが重要な時間的成分を持つという考えは、既存の解析では必ずしも適用されず、代わりに、プリミティブな脊椎の相乗効果は純粋に空間的なものであり、より高いレベルの制御によって時間的成分が追加されると仮定されています。たとえば、これはマカクザルの運動皮質からの筋肉の相乗効果の分析で観察されており、時間の経過とともに活性化されるより基本的な空間パターンの組み合わせから時空間パターンを構築することができます ([[@OverduindAvellaRohEtAl15]])。同様に、[[@^BergerMasciulloMolinariEtAl20]] は、小脳が基本的な空間プリミティブに時間的要素を追加することを示しました。
それにもかかわらず、これらの原則の両方を同時に採用することは可能です(つまり、プリミティブ内の時間的ダイナミクスと、より高いレベルの制御によって課される追加の時間的構造)が、そうすることで、高次元EMG(筋肉から記録された筋電図信号)または他の同様のデータから筋肉の相乗効果を抽出する標準的な分析アプローチに混乱が生じます。
{id=”図_脳幹” スタイル=”高さ:40em”}
大まかなレベルでは、追加の各制御レベルの寄与は次のとおりです ([[@GrillnerElManira20]]、[[@ArberCosta22]]、[[@ZaaimiDeanBaker18]])。
-
脳幹核は、より広範な感覚入力を統合し、脊椎の相乗効果の組み合わせとして、より複雑で生存に関連した行動、たとえば、運動活動の制御 ([[#figure_brainstem]]) や食物に手を伸ばす動作 ([[@EspositoCapelliArber14]]; [[@CapelliPivettaSoledadEspositoEtAl17]]; [[@ZeleninOrlovskyDeliagina07]]; [[@ZaaimiDeanBaker18]]) を駆動します。
-
[[cerebellum]] は 2 つの異なるタイプの予測学習を実行し、脊椎シナジー自体の直接調整 ([[@UdoMatsukawaKameiEtAl80]]; [[@GrillnerElManira20]]; [[#figure_cerebellum-tuning]]) と、複数の脊椎シナジーにわたって異なる調整された時空間パターンを学習する機能 ([[@BergerGentnerEdmundsEtAl13]]) を実行します。小脳を通るさまざまな経路(前庭脊髄、赤核脊髄、網状脊髄)で行われる機能。
-
[[basal ganglia]] の背外側 (運動) 領域は、[[dopamine]] 駆動の [[reinforcement learning]] メカニズムに基づいて進行中の運動動作を動的に調整し、報酬につながる動作の形成と選択を最適化し、マイナスの結果を回避します ([[@GrillnerRobertsonKotaleski20]]; [[@ParkCoddingtonDudman20]]; [[@ArberCosta22]]; [[@MarkowitzGillisBeronEtAl18]])。
-
哺乳類、特に霊長類では、前頭葉の [[neocortex]] は脊髄原始のよりきめの細かい時空間調節 ([[@OverduindAvellaRohEtAl15]]; [[@AflaloGraziano06]]) を提供し、霊長類の外では不可能な方法で個々の運動単位を直接制御する機能さえ備えています。これはおそらく、霊長類、特に人間が道具を使用するための、新しく高度な運動プログラムを開発する能力の基礎となっていると考えられます ([[@MarshallGlaserTrautmannEtAl22]]; [[@StrickDumRathelot21]])。
より抽象的な計算レベルの分析では、[[@^Brooks86]] によって明確化された [[subsumption]] アーキテクチャが、これらの上位レベルの制御スタックがどのように機能するかについて影響力のあるフレームワークを提供します。レベルを追加するたびに、新しい「上位」機能が追加され、既存の下位レベルの機能の上に_構築_されます。下位レベルのシステムはそのまま残り、システムはいつでもそれに頼ることができますが、上位レベルのシステムが関与する必要がある場合、制御を奪うために競合する下位レベルのシステムを選択的に禁止することができます。全体的な動作は、さまざまなレベルにわたる多数の分散制御システム間の並列相互作用から生じる [[emergent]] ダイナミックとして想定されます。
PCT フレームワークもコントローラーの階層システムを想定していますが、特に、上位レベルの制御システムは下位レベルのシステムの制御パラメーターを設定することによって機能すると想定しています。これは、低レベルの阻害とは異なります。
最適フィードバック制御 (OFC) は、利用可能なフィードバック信号を使用してモーター パラメーターを適応的に制御できる最適なオンライン コントローラー メカニズムを確立することにより、質的に異なる制御形式を提供します。このコントローラーには、小脳が提供すると広く信じられている内部順モデルが必要です。 OFC 自体は抽象的な計算レベルで定式化されているため、必ずしもこれらの神経クレームに基づいているわけではありません。それにもかかわらず、脊髄レベルの筋肉の相乗効果を調整し、適応制御の他の側面を推進する、ある種の高次レベルの制御の能力は、平衡点制御と OFC ([[@Shadmehr10]]) の重要な違いを表しています。
UMC (非制御多様体) フレームワークは、一見冗長な制御パラメータ ([[@GelfandLatash98]]; [[@LatashScholzSchoner07]]) の多様体全体を持つことによってもたらされる柔軟性と適応性を強調しています。この柔軟性は、小脳で学習された予測順モデルに基づいて構築された、大脳基底核、特に新皮質における最高レベルの制御によるものと考えられます。
これらのさまざまな形式の階層制御は相互に排他的ではなく、複数レベルの制御の存在と一致しており、入手可能なかなりの証拠があります。ただし、ワールド モデル コントローラーの概念など、よりモノリシックな形式のモーター制御とは重要な対照があります (詳細については、[[reinforcement learning]] を参照)。
残りのセクションでは、筋肉から始まり、そこから脊髄、脳幹、小脳、大脳基底核、新皮質に至るまで制御レベルを上げていく、骨格運動システムの基本的な特性について説明します。
筋肉
{id=”figure_muscle” style=”高さ:30em”}
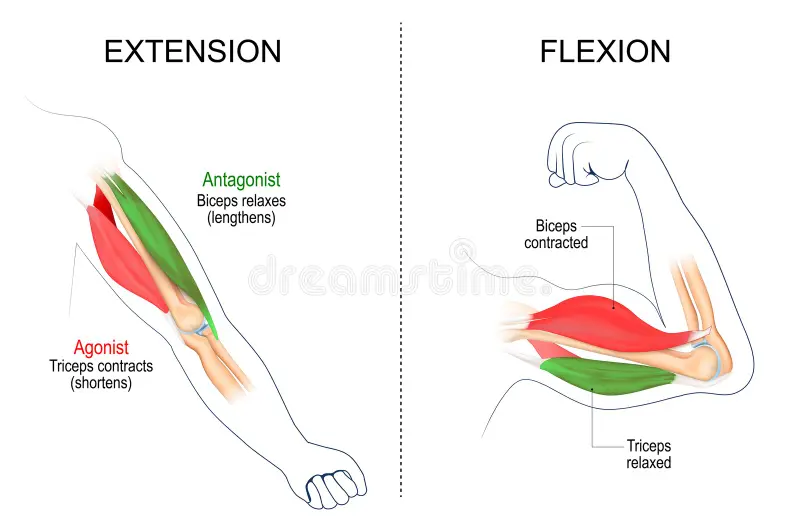
骨格筋は [[opponent]] ペア ([[#figure_muscle]]) に編成されており、伸筋の収縮により四肢が外側に拡張され (extension)、屈筋の収縮により四肢が内側に収縮されます (flexion)。筋肉は、腱を介して骨に接続されています。腱は、コラーゲンでできた強力でわずかに弾性のあるロープ状の組織です。腱は通常、関節の周囲を包み込み、より大きなてこの作用を得るために骨のさらに下で接続することによって力を伝達します。
筋肉は、運動ニューロンからの [[acetylcholine]] の放出によって駆動される収縮によってのみ力を発揮し、アクチン線維とミオシン線維の収縮を引き起こします。したがって、実際に手足を動かすには、片側が収縮し、もう一方が弛緩するように、両側の相手の筋肉を調整する必要があります (筋肉が活性化されていない場合、これは自然に起こります)。さらに、筋肉全体 (例: 上腕二頭筋) は、サイズが異なる個々の 運動単位 で構成されており、それぞれが独自の運動ニューロンを持っています。
これらの異なる運動ユニットは通常、「サイズ原理」 ([[@Henneman85]]) に従って活性化されます。この原理では、下降する運動制御信号が最初に小さな力を及ぼす小さなユニットを動員し、その後、発火率と活性化された求心性入力の数が増加するにつれて、ますます大きなユニットが続きます ([[@LucaErim94]])。これは、小型ユニットの方がしきい値が低く、大型ユニットよりも発砲率が高いためです。このサイズの原則により、疲労効果も最適化されます。
サイズ原理により、単一の共通駆動一次元コマンド入力が、特定の筋肉全体のすべての運動単位に投影されることが可能になり、この 1 つの入力により、神経活動の増加に伴って収縮力も自然に増加するようになります。これは、モーター システムが制御問題の有効次元を削減するもう 1 つの方法です。
眼筋
{id=”figure_ocular-muscles” style=”高さ:40em”}
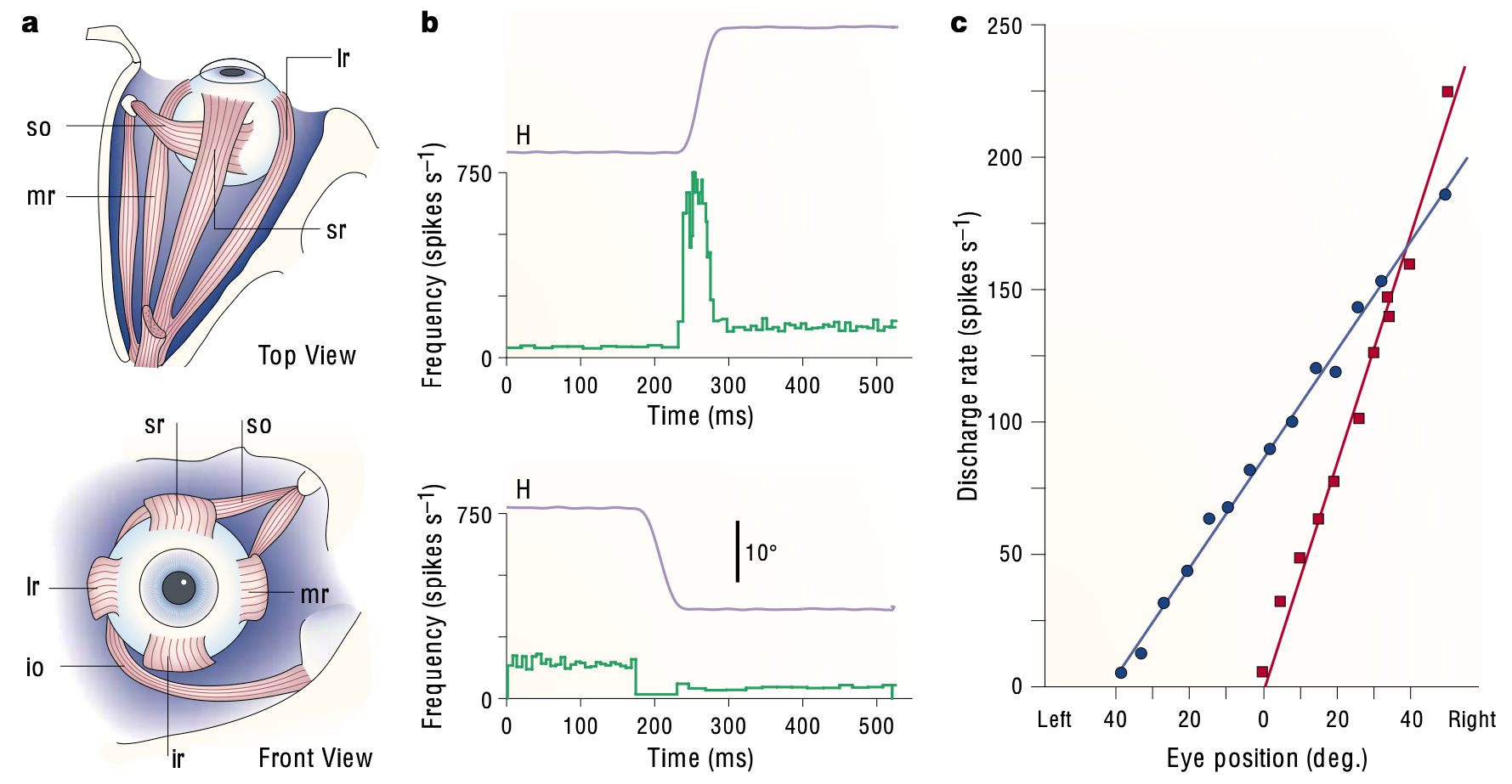
霊長類の目の位置を制御する筋肉は特によく研究されており ([[@Sparks02]]、[[@SylvestreCullen99]]、[[@FuchsScudderKaneko88]])、[[@^Sparks02]] から [[#figure_ocular-muscles]] に示すように、運動制御の比較的単純な例を提供します。目の位置と目の速度と、水平方向の目の動きを制御する外転核ニューロンの正味の発火との間には直接的な線形関係があります。目を中心の位置から外すにはさまざまなレベルの筋肉の収縮が必要となるため、これらのニューロンの持続発火率によって目の位置が決まります。大きな位相バースト (最大 750 Hz、つまり 1.3 ミリ秒ごとのスパイク!) は目の急速な動きを引き起こします。これは目の回転速度に直線的に関係し、バースト全体の持続時間は目の動きの持続時間と相関します。
{id=”figure_ocular-brainstem” style=”高さ:45em”}
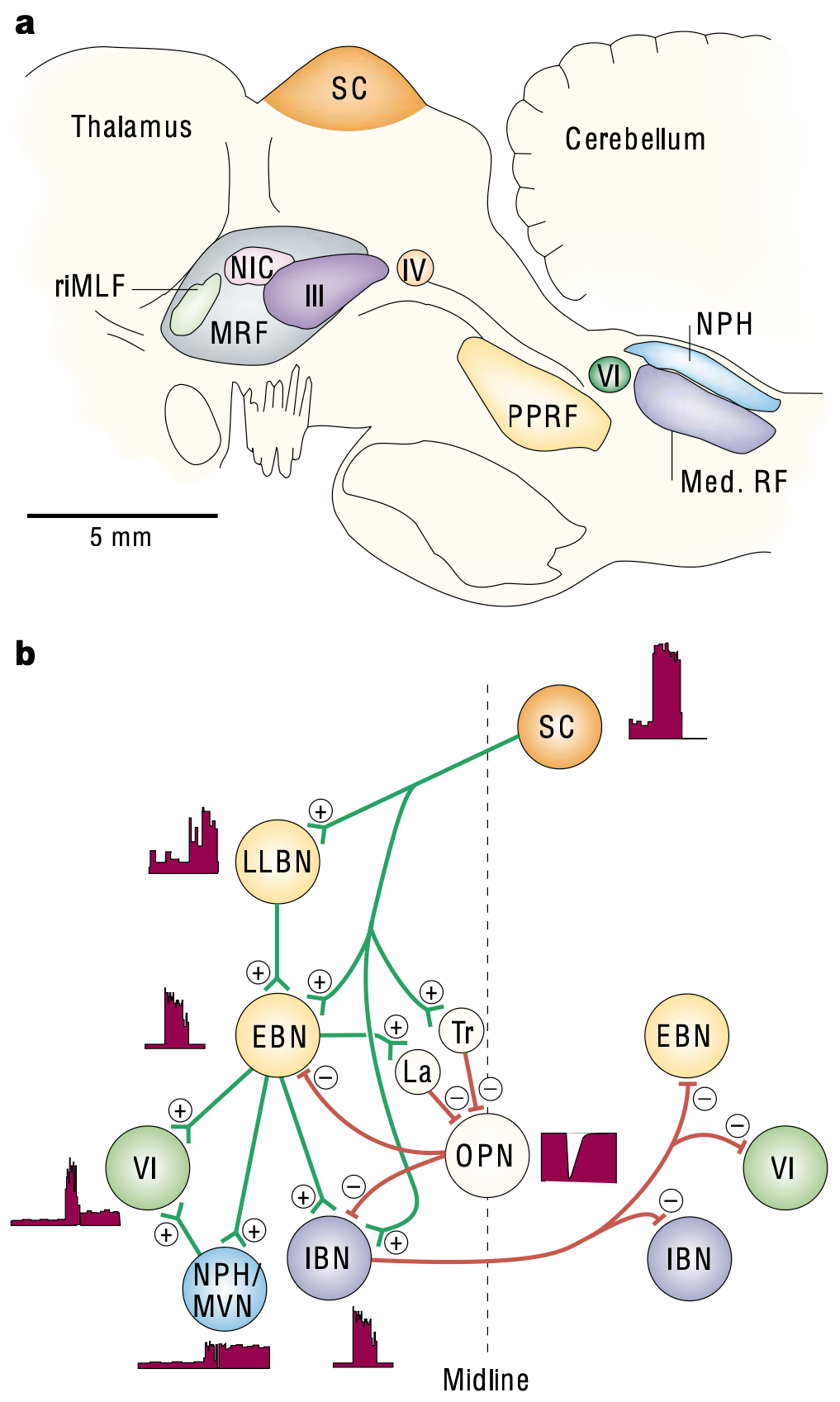
図 [[#figure_ocular-brainstem]] は、外転ニューロンで観察される活動パターンの生成に関与する主要な脳幹領域の一部を示しています。傍正中橋網様体 (PPRF) と髄様網様体 (Med RF) には、図に示すようにさまざまなタイプのバーストおよび休止ニューロンが含まれており、これらがサッカードに関連する強力な位相性バーストを駆動します ([[@Sparks02]])。 [[vestibular]] 二次核 (NPH および MVN) のニューロンは、現在の目の位置に比例して持続的な固視維持発火を駆動します。
これらのネットワークは脊髄筋の相乗効果に相当しますが、目は体幹ではなく頭にあるため、脳幹に直接位置しています。運動ニューロンの発火のより複雑で高次元のパターンが運動筋、到達筋、およびその他の骨格筋の領域で必要とされるのと比較して、目の運動出力の特に単純な性質により、このシステムは比較的容易に理解できます。それにもかかわらず、必要な時間的パターンを生成するために必要なニューロン発火パターンの多様性という点では、おそらく同じ種類の原理が当てはまると考えられます。
[[superior colliculus]] (SC) は、これらの脳幹眼球運動野 ([[@IsaMarquez-LegorretaGrillnerEtAl21]]) に主な入力を提供します。これは、聴覚およびその他のモダリティからの感覚信号を統合し、視覚的な眼球運動(サッカード)の空間的標的化を駆動するニューロンの地形的網膜マップを備えています。運動制御の階層的な [[subsumption]] の性質と一致して、一次運動野の前頭眼野 (FEF) ニューロンは、通常、SC への入力を通じて作用し、視覚サッカードのより柔軟なターゲティングを駆動しますが、中脳回路を直接駆動することもできます ([[@Sparks02]])。
丘式筋肉モデル
{id=”figure_hill-muscle” style=”高さ:25em”}
![筋肉自体を表す収縮要素 (CE)、筋肉の軟組織のバネのような特性を表す平行な受動要素 (PE)、および腱を表す直列のバネ状要素 (SE) を備えた丘型筋肉モデル。 [ウィキメディア・コモンズ] から転載](https://commons.wikimedia.org/wiki/File:Hill_muscle_model.svg) .](media/fig_motor_hill_muscle.png)
.](media/fig_motor_hill_muscle.png)
筋肉のシミュレーションに使用される標準方程式は、もともと [[@^Hill38]] によって開発された単純化された近似に基づいており、したがってヒル型筋肉として知られています ([[@RitchieWilkie58]]; [[@Zajac89]]; [[@MillardUchidaSethEtAl13]]; Millard (2013) バージョンの実装については OpenSim を参照してください: [[@SethHicksUchidaEtAl18]]; ギットハブ)。この筋肉モデルには、並行して動作する 2 つの要素 ([[#figure_hill-muscle]]) があります。1 つは収縮する実際の筋肉を表す contractile 要素 (CE)、もう 1 つは CE と平行に実行され、筋肉の軟部組織のバネのような特性を表す passive 要素 (PE) です。腱は、筋肉 CE と直列 (SE) のバネ状要素を介して表されます。
[[@^MillardUchidaSethEtAl13]] は、この基本的な筋肉モデルのさまざまなバリアントの詳細な比較を提供し、バネ状の腱 (SE) を備えた完全なモデルの減衰バージョンが精度と計算効率の間の最良の妥協点を提供することを示しています。彼らはまた、剛腱を備えたバージョンも分析しました。これは計算がより単純で高速ですが、かなり不正確でもあります。
{id=”eq_hill” title=”ダンプヒル型モデル”} \(F_0 (a f_L(l) f_V(v) + f_{PE}(l) + \beta v) cos \alpha - F_0 f_T(l_T) = 0\)
ここで、$F_0$ はピーク等尺性力、$a$ は正規化された活性化レベル、$l$ は現在の正規化された筋肉の長さ、$v$ は速度、$f_L$ は力の長さ関数、$f_V$ は力-速度関数、 $f_{PE}$ は受動的力-長さ曲線、$\beta$ は減衰係数 (デフォルトは 0.1)、$f_T$ は腱の力-長さ関数、$\alpha$ は腱に対する筋肉のペネーション角度です。これらのパラメーターと機能はすべて解剖学的に導き出すことができますが、筋肉によって異なるため、このレベルでのモデリングは通常、より生物学的な情報に基づいた方法で行われます。
方程式 [[#eq_hill]] は、離散時間シミュレーションで筋肉の長さを反復的に更新するために、ニュートン法などの微分ベースの根探索法を使用して速度 $v$ を求めることができますが、これはある程度の計算量を要します。
別の解決策は、$f_V(v)$ の区分線形近似 ($v=0$ で傾きが大幅に変化しますが、それ以外は比較的線形です) を使用して、$v$ を解析的に解決できるようにすることです。
{id=”eq_hill-about” title=”ダンプ ヒル タイプ モデル”} \(v = \frac{f_T(l_T) sec(\alpha) - f_{PE}(l)}{a f_L(l) f_V + \beta}\) ここで、$f_V$ は、$v$ の指定された値に関連する傾きを表します ($v > 0$ かどうかに応じて、それに収束するには 2 回の反復が必要です)。
筋肉をシミュレートするさらに簡単で精度は低い方法は、コージェントコアラボの物理学 モデルで行うように、標準の物理シミュレータ ジョイントの剛性と減衰パラメータを操作しながら、ジョイントの位置の値を更新することです。これには、(平衡点制御の場合と同様に) 目標の関節位置を知る必要がありますが、剛性パラメーターは関節の動きの速度に比例し、減衰によって減衰されたバネのダイナミクスが提供されます。
## 脊髄
{id=”figure_spinal-pathways” style=”高さ:35em”}
![脊髄内の主要な経路。上行性感覚経路は後角灰白質のニューロン(紡錘線維の筋長とゴルジ腱の伸張固有受容、接触、痛み)に由来し、前角には下行性の運動入力を受け取る運動ニューロンが含まれており、また上行遠心性コピー信号も送信します。中間灰白質には副感覚ニューロンと運動ニューロンが含まれており、介在ニューロンが存在し、これらが実際に下降制御入力のほとんどの主なターゲットとなります。 [ウィキメディア・コモンズ] から転載](https://commons.wikimedia.org/wiki/File:Spinal_cord_tracts_-_English.svg) .](media/fig_spinal_cord_pathways.png)
.](media/fig_spinal_cord_pathways.png)
{id=”figure_spinal-interneurons” style=”height:45em”}
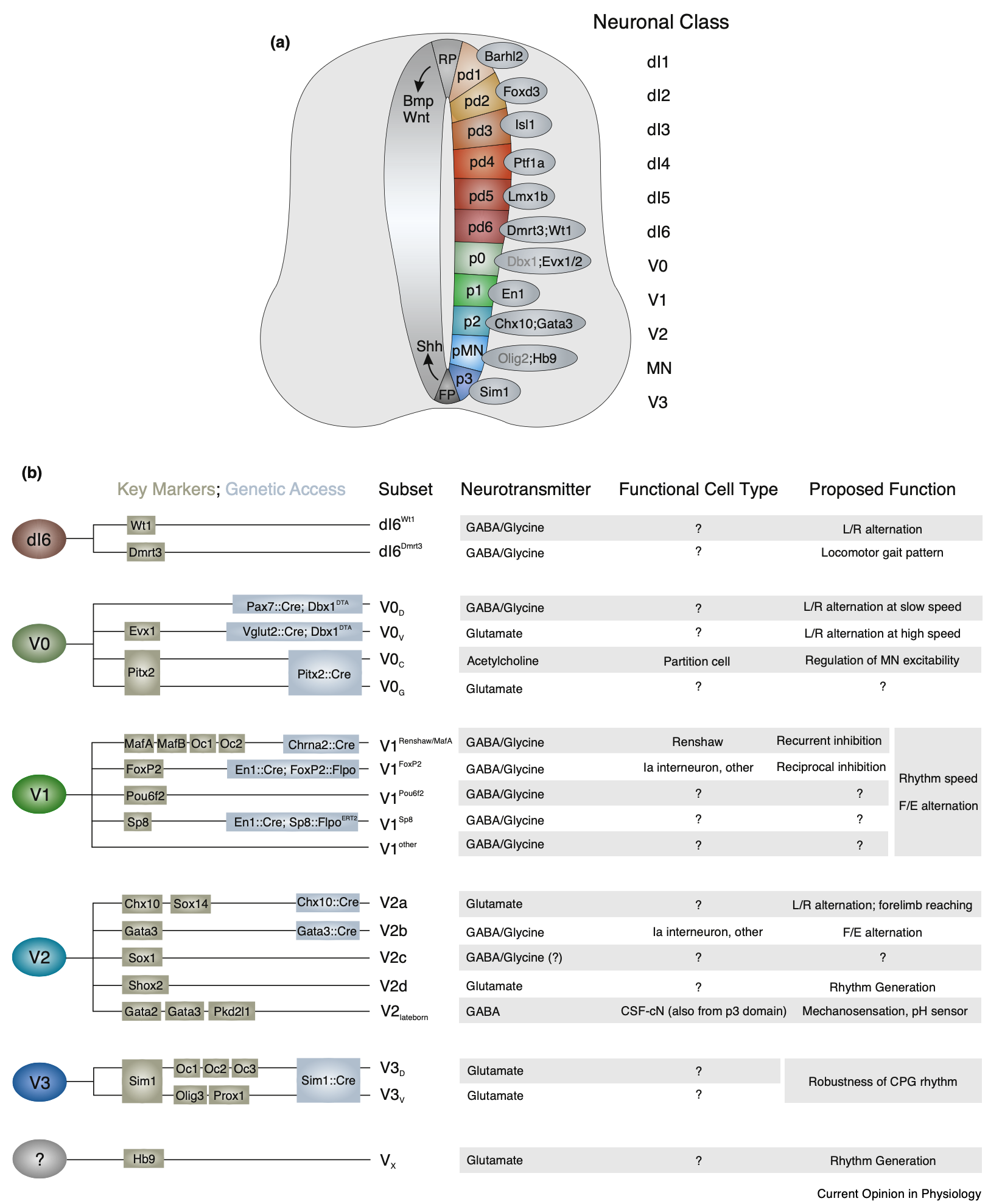
脊髄は、脊髄の広範な神経ネットワークによって直接最下位レベルで実現される「筋肉の相乗効果」に関して上で説明したように、受動的なモーター出力システムからはほど遠いものです。これらのニューラル ネットワークには、一次感覚ニューロンと運動ニューロン、および接続と神経伝達物質の異なるパターンを持つ多くの異なるサブタイプの介在ニューロンが含まれます ([[#figure_spinal-interneurons]] から [[@Bikoff19]]、[[@SenguptaBagnall23]] も参照)。
このシステムの入力と出力は、[[#figure_spinal-pathways]] ([[@TanFaullCurtis23]]、[[@WatsonKayalioglu09]] も参照) に示されているように、長距離軸索線維のさまざまな経路に組織化されており、後角 灰白質には、さまざまな体細胞入力に応答する感覚ニューロンの細胞体があり、その入力は上行路を通って身体の多くの領域に伝達されます。脳、最後に [[thalamus]] と体性感覚 [[neocortex]] を含みます。これらの体性感覚信号には次のものが含まれます。
- 皮膚および皮下領域の機械受容体からの接触、振動、圧力、および張力の信号。
- 侵害受容体からの痛みと温度。
- 筋肉の長さ (Ia 型感覚ニューロンを介して) および筋肉全体の長さ (II 型ニューロンを介して) の変化に応答する_筋紡錘_ 線維を介した固有受容信号、および四肢の伸張または負荷に応答する_ゴルジ腱受容体_ (Ib 型ニューロンを介して) を介した固有受容信号。
前角灰白質には筋肉の収縮を直接引き起こす一次運動ニューロンが含まれており、中間灰白質領域(脊髄の特定のレベル)には感覚ニューロンと運動副ニューロンが含まれています。
介在ニューロンの広範なネットワークは、下降運動コマンド入力と局所 (およびより遠位) 感覚入力を直接受け取り、運動動作の低次元語彙を生み出す筋肉の相乗効果を実現します。感覚信号 (特に筋紡錘長信号) を直接組み込むことにより、これらの筋肉の相乗効果コンポーネントが堅牢かつ適応的になり、下降運動制御入力の仕事が簡素化されます。
これらの介在ニューロンは以前はいくつかの特定のクラス ([[@McCrea92]]; [[@Hultborn06]]; [[@CoteMurrayKnikou18]]) に分類されていましたが、[[#figure_spinal-interneurons]] ([[@Bikoff19]]; [[@Bikoff19]]; [[@SenguptaBagnall23]])。重要なことに、感覚入力は複数の異なる介在ニューロンに投影され、各介在ニューロンは複数の異なるタイプの入力を受け取ることができるため、歴史的な説明とは異なり、すべてがはるかに複雑で分類するのが困難です([[@Jankowska22]]; [[@CoteMurrayKnikou18]])。
{id=”figure_spinal-cpg” style=”高さ:45em”}
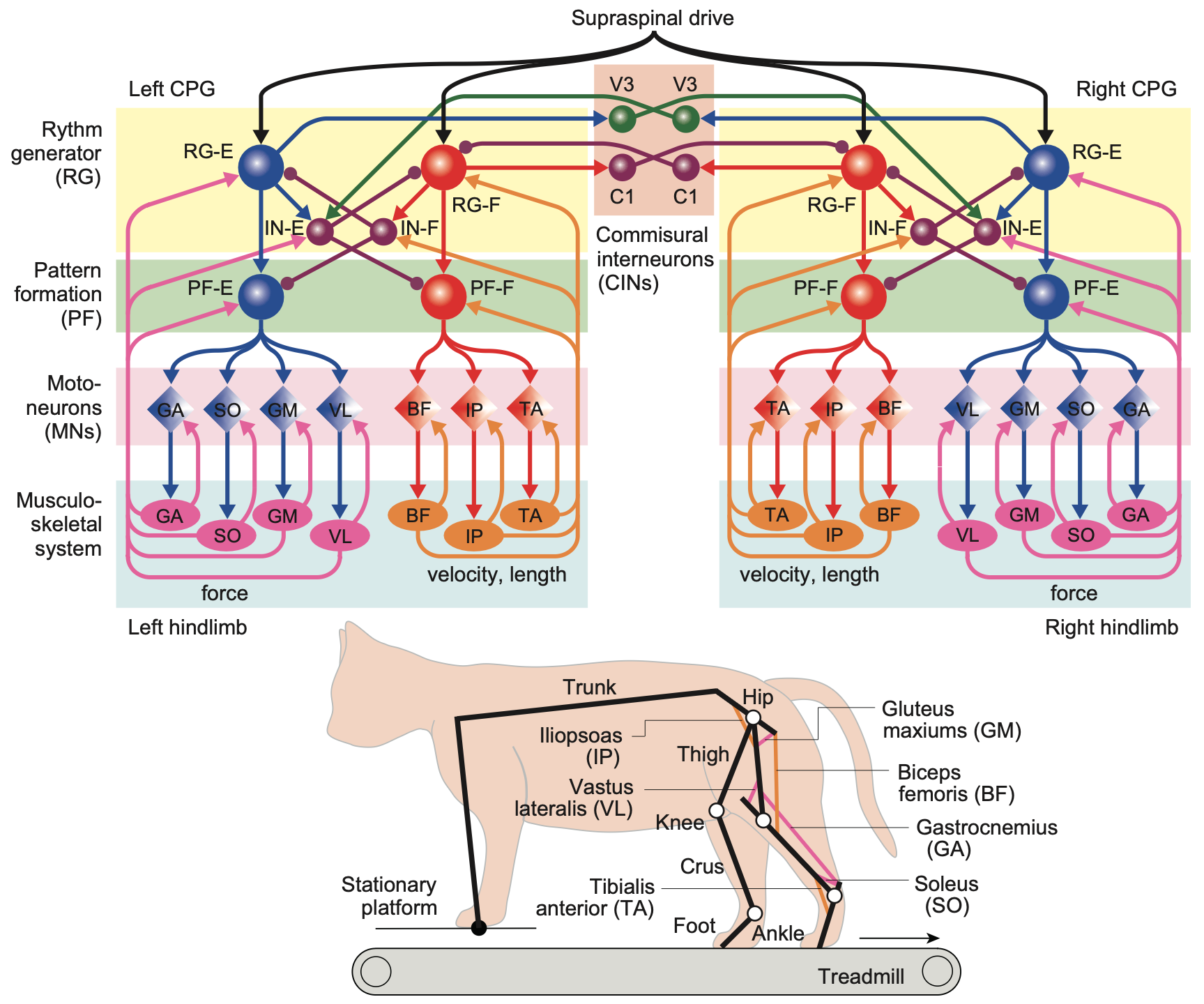
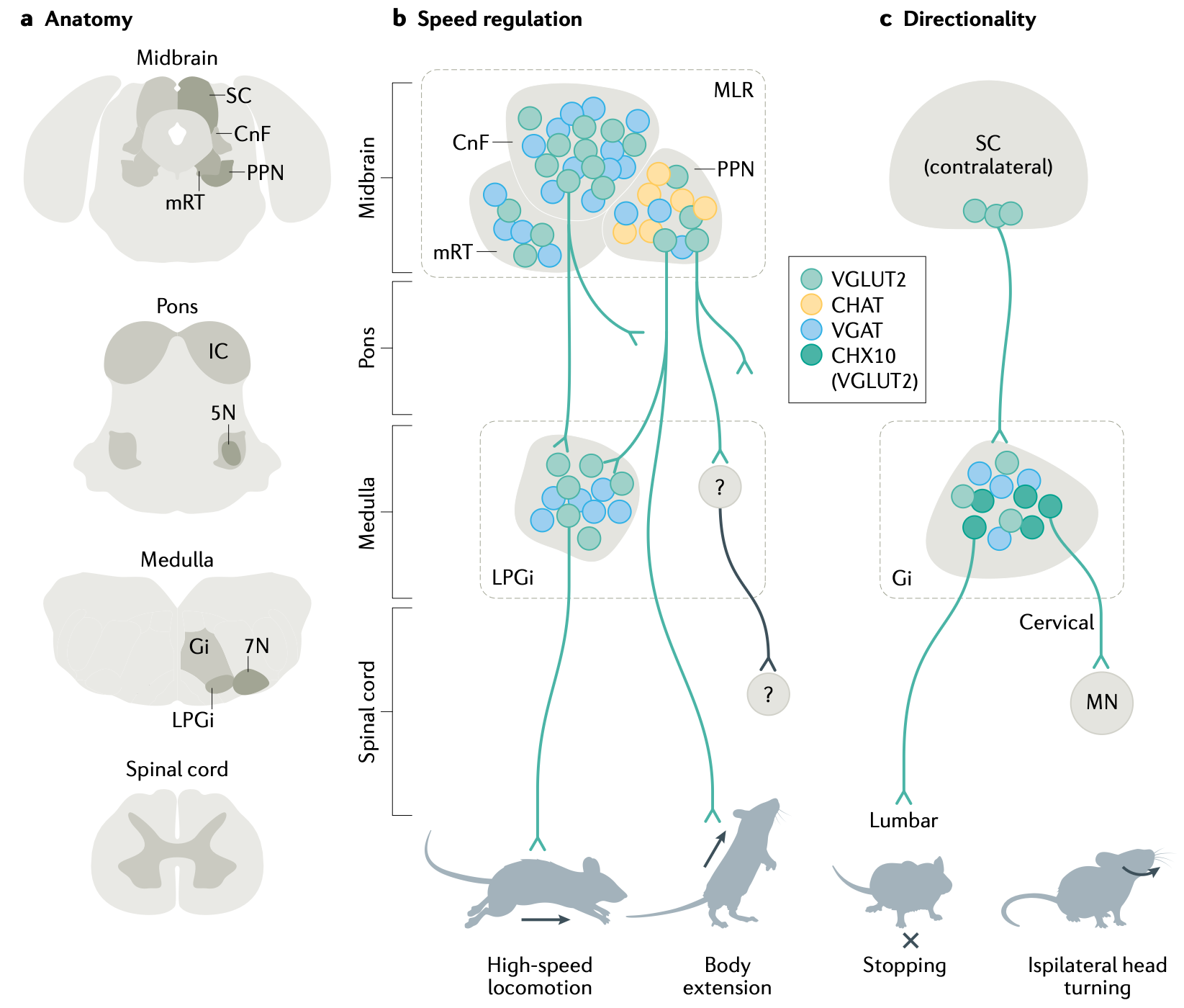
これらの脊椎回路の高度さと複雑さの最近の例は、[[@^ShinoharaAmbeKimEtAl25]] から [[#figure_spinal-cpg]] に示されています (関連モデルのレビューについては [[@DannerShevtsovaFrigonEtAl17]] を、人間の二足歩行の例については [[@AoiOhashiBambaEtAl19]] を参照してください)。これは、サポートのない「穴」に足が着地するときなど、後肢の移動動作の多くの側面をシミュレートできます。この堅牢な適応動作は、皮膚の接触入力とともに、筋紡錘線維からの固有受容長および速度信号が回路全体に直接組み込まれた結果として生じます。全体的なエネルギー消費を最小限に抑え、凹凸のある表面に適応するように接続が最適化され、そこからサポートが不足している状況に自動的に一般化されました。
重要なことに、これらの堅牢な適応行動には、半球全体にわたる両側の調整が必要です。これは、一方の足がサポートされていない場合、安全な足場が得られるまで、もう一方の足が荷重を負担しなければならないためです。このモデルは、対応する実験準備における運動と神経発火信号の詳細なパターンを予測でき、これらの回路がどのように機能するかについての強力に検証された理解を提供します。
興味深いことに、これらの回路には、下降入力制御信号の発火率の関数として移動速度を調整するという特性もあり、この複雑な調整された運動動作のすべてを実行する方法を学習する必要があるという見通しに比べて、高次脳領域の仕事がはるかに簡単になります。
これらの CPG ダイナミクスを理解するためのより抽象的な分析アプローチでは、状態空間法を使用して、運動に関与する脊髄介在ニューロンの集団全体にわたるダイナミクスを特徴付けました ([[@WimalasenaPandarinathAuYong25]])。このアプローチでは、特定された個々の介在ニューロンの詳細な動作を説明する機能は省略されますが、完全な集団動態を組み込むことができます。
霊長類と人間はより多くのコントロールを持っています
霊長類、特にヒトは、筋肉の相乗効果や、脊髄と脳幹のネットワークによって実現されるCPG回路によって明らかにされるものを超えて、個々の筋肉を直接制御するはるかに優れた能力を持っているというかなりの証拠があります([[@GrillnerElManira20]]; [[@Grillner81]])。たとえば、[[@^MarshallGlaserTrautmannEtAl22]] は、上で議論したサイズ原理の厳格な制約を超えた、運動単位のより柔軟な動員の証拠を示しました。また、霊長類には、特定の四肢へのより集中的な特殊な投影を有する二次運動野が多数存在します ([[@StrickDumRathelot21]])。
興味深いことに、このきめ細かな制御レベルの増加に伴う追加の自由度という考えにもかかわらず、脳の大きさと、ヒトや他の霊長類が依然として従っている初移動年齢との間には信頼できる関係がある([[@GarwiczChristenssonPsouni09]])。
[[@^Bernstein96]] は、人間の運動制御の発達は、より厳格で進化的にコード化された運動プログラムへの依存から、徐々に新皮質からのより柔軟で高次元の制御の学習を可能にする軌道をたどると仮説を立てました。証拠はこの仮説と一致しています ([[@HinnekensBarbu-RothDoEtAl23]]; [[@DominiciIvanenkoCappelliniEtAl11]])。
より高いレベルのシステム
残りのセクションでは、より高いレベルの制御を提供する神経システムに関連する問題のいくつかをさらに詳しく検討します。これらのシステムに関する最近の広範な解説については、[[@^BizziAjemian20]] を参照してください。
小脳
{id=”figure_小脳チューニング” style=”高さ:30em”}
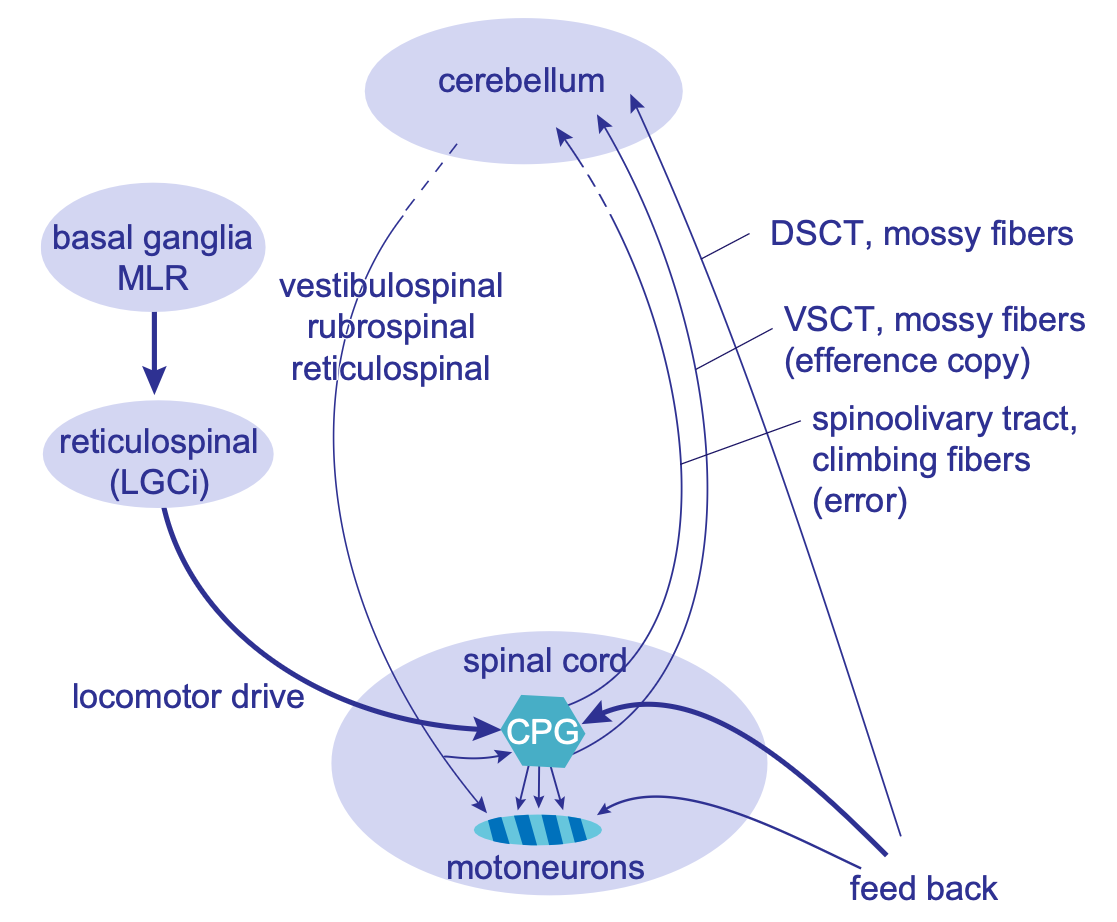
[[cerebellum]] は、他の進行中の感覚運動信号の広範囲を使用して、特定の感覚信号を予測することによって学習します。この予測学習を使用して、予想される信号を適応的にフィルター処理し、これらの信号の早期のアクティブな予測を順方向モデルとして提供することもできます。これらの学習能力は、[[#figure_cerebellum-tuning]] から [[@^GrillnerElManira20]] に示すように、脊椎 CPG と筋肉の相乗効果の微調整を促進するために使用できます。
小脳は、予測学習を使用して筋肉の相乗効果やその他のより高度な運動プログラムを活性化し、より適切に調整された運動動作を達成することもできます。よく研究された例の 1 つは、[[cerebellum#vestibulo-ocular reflex]] (VOR) の場合です。この場合、頭の回転の予期される影響により、予期的、代償的な眼球運動が反対方向に駆動され、安定した視覚知覚が維持されます。
ただし、場合によっては、代わりに、頭と目の動きを組み合わせて、空間内の特定の点を注視するサッカードを生成することが目標である場合もあります。この場合、VOR 反射を抑制し、予期される頭の動きを考慮した別の衝動性眼球運動をプログラムする必要があるため、2 つの動作が一緒になって最終的に目が目標位置に固着することになります。したがって、VOR 対サッケードのこの柔軟な活動空間は、小脳における高次の制御がどのように機能するかについてよく研究された良い例を表しています。
小脳はさまざまな運動プログラムの一連の制御「ノブ」を_管理_しますが、小脳自体がこれらのノブをどうするかを_決定_するわけではないことを理解することが重要です。これは大脳基底核の仕事であり、大脳基底核はそのような決定を下すための報酬に基づく学習メカニズムを備えています。その代わりに、小脳の学習は、運動動作の影響を予測し、その予測に従ってすべてを駆動する能力を活用することで、さまざまなプログラム間でスムーズに調整できるように最適化されます。
大脳基底核
{id=”figure_bg-outs” style=”高さ:30em”}
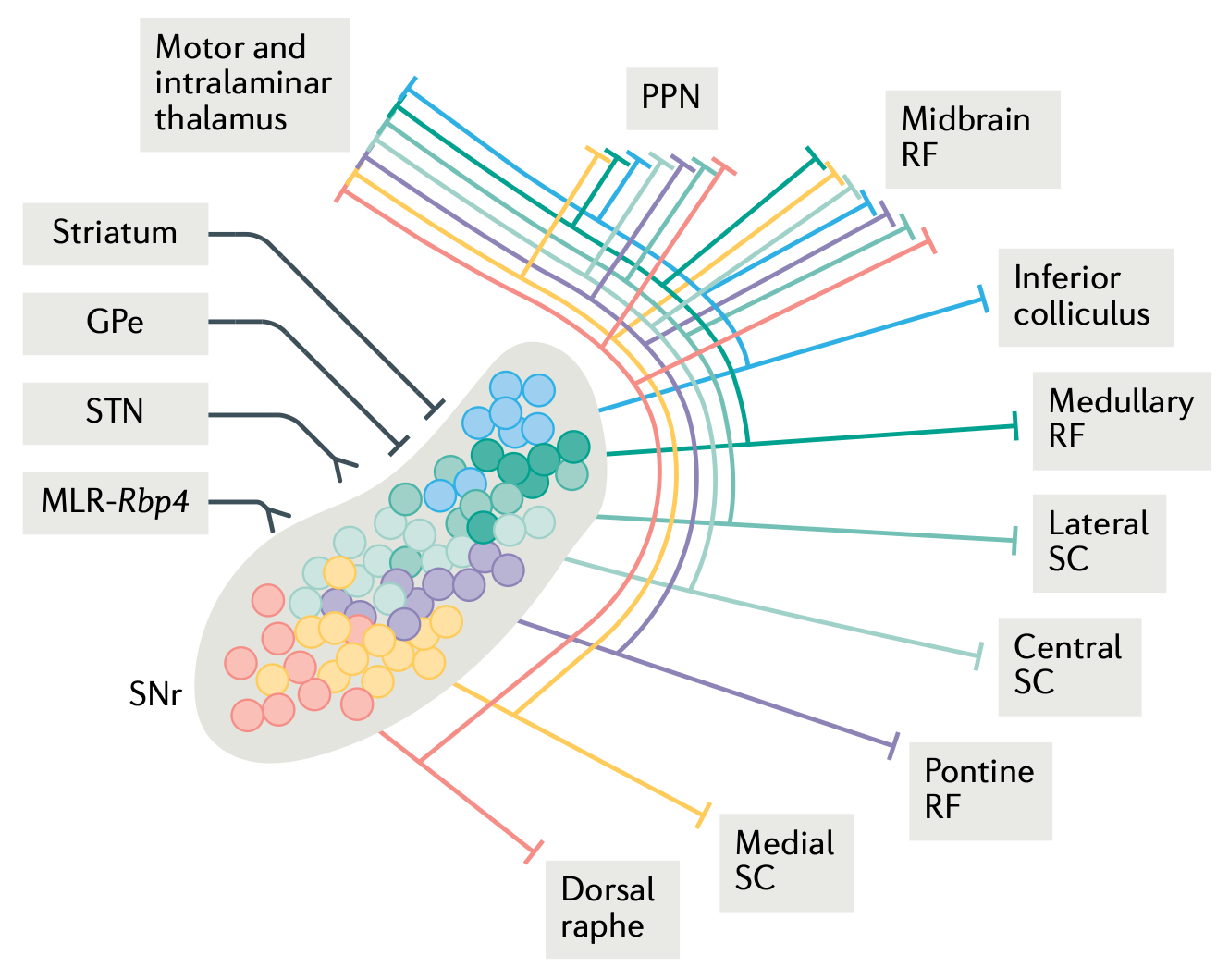
背外側 (DL) [[basal ganglia]] (BG) は、関連するすべての下位レベルの運動制御領域 ([[#figure_bg-outs]]、[[@ArberCosta22]] から) に投影される広範囲の出力ニューロン (SNr および GPi 内) の学習を促進します。 [[basal-ganglia#Motor activity in DL]] で説明したように、BG は下降するモーター信号の動的で連続的な変調を提供します。これにより、強力な勾配ベースの [[search]] による学習の推進が可能になります。これは、どの形式の離散的な Go vs. NoGo モーター選択ロジックよりもはるかに効率的です。
| BG はまた、さまざまな [[thalamus | thalamic]] 経路を介して前頭皮質に上行入力を送りますが、これらの経路の拡散した性質により ([[#figure_bg-outs]] に示されている SNr からのはるかに特殊化された集中的な出力と比較して)、これらの上行信号の機能が不明瞭になっています。次のセクションでは、いくつかの可能性について説明します。 |
前頭葉
{id=”figure_homunculus” style=”高さ:35em”}
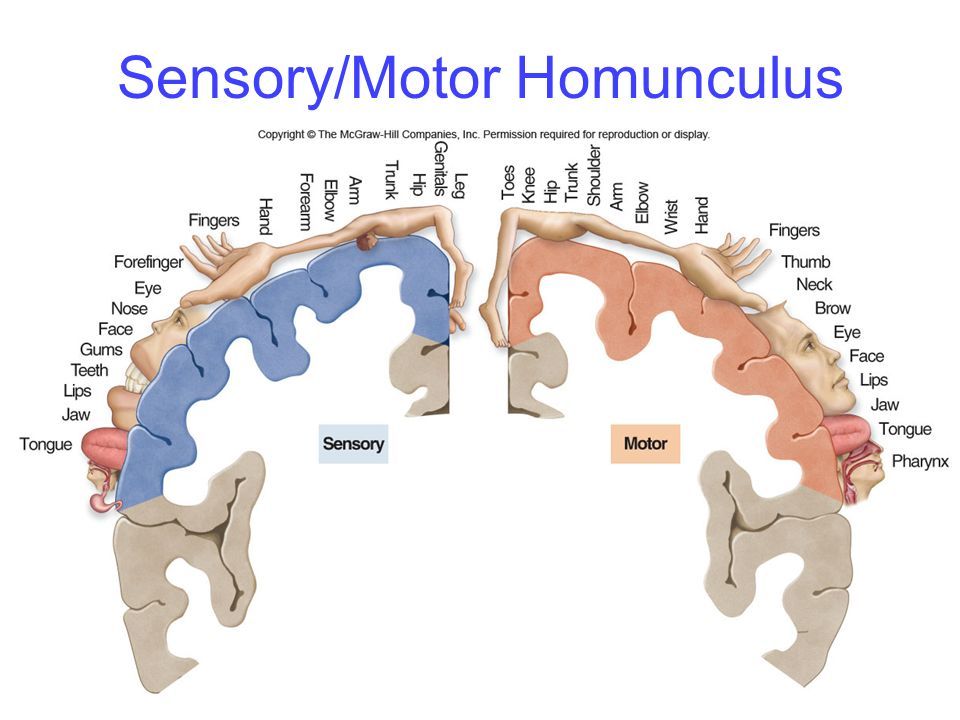
[[neocortex]] は、(哺乳類における) 最高レベルの運動制御であり、すべての運動信号と体性感覚信号の広範なマッピングが、脳内の一種の「ホムンクルス」 (「小人」) として説明される「体性トピック」マップに編成されています ([[#figure_homunculus]])。霊長類、特にヒトにおける新皮質の重要性の増大と一致して、これらの体性感覚および運動表現に充てられる大量の神経組織により、人々は最も柔軟で多様な形の運動行動、たとえば数年ごとのオリンピックで展示されるあらゆるものを示すことが可能になります。
脳の他の部分とは異なり、新皮質は、動物の一生の間に行われる学習を通じて、その機能のほぼすべてを獲得すると考えられています。これは運動システムに重要な課題を投げかけています。運動皮質領域は脊髄の「言語を話す」ことをどのように学習し、意味のある運動制御信号を提供できるのでしょうか?
私たちの仮説は、[[thalamus]] の肺核と MD 核によってサポートされている、大脳皮質の感覚表現やその他の表現の [[predictive learning]] で使用されているのと同じメカニズムが、前頭皮質領域の運動側でも働いている可能性があるというものです。 [[thalamus#frontal thalamus]] で説明したように、VL 野 (腹外側) は小脳核からドライバー入力を受け取り、より焦点の中心のような入力を運動皮質の中層に送ります。これらの入力は、運動皮質に対する元のコマンドと、運動システムがそのコマンドで実際に行ったことを反映する後続の遠心性コピーとの間の時間的差異を反映する適切な運動コマンドの学習を促進するのに十分に特異的である必要があります。
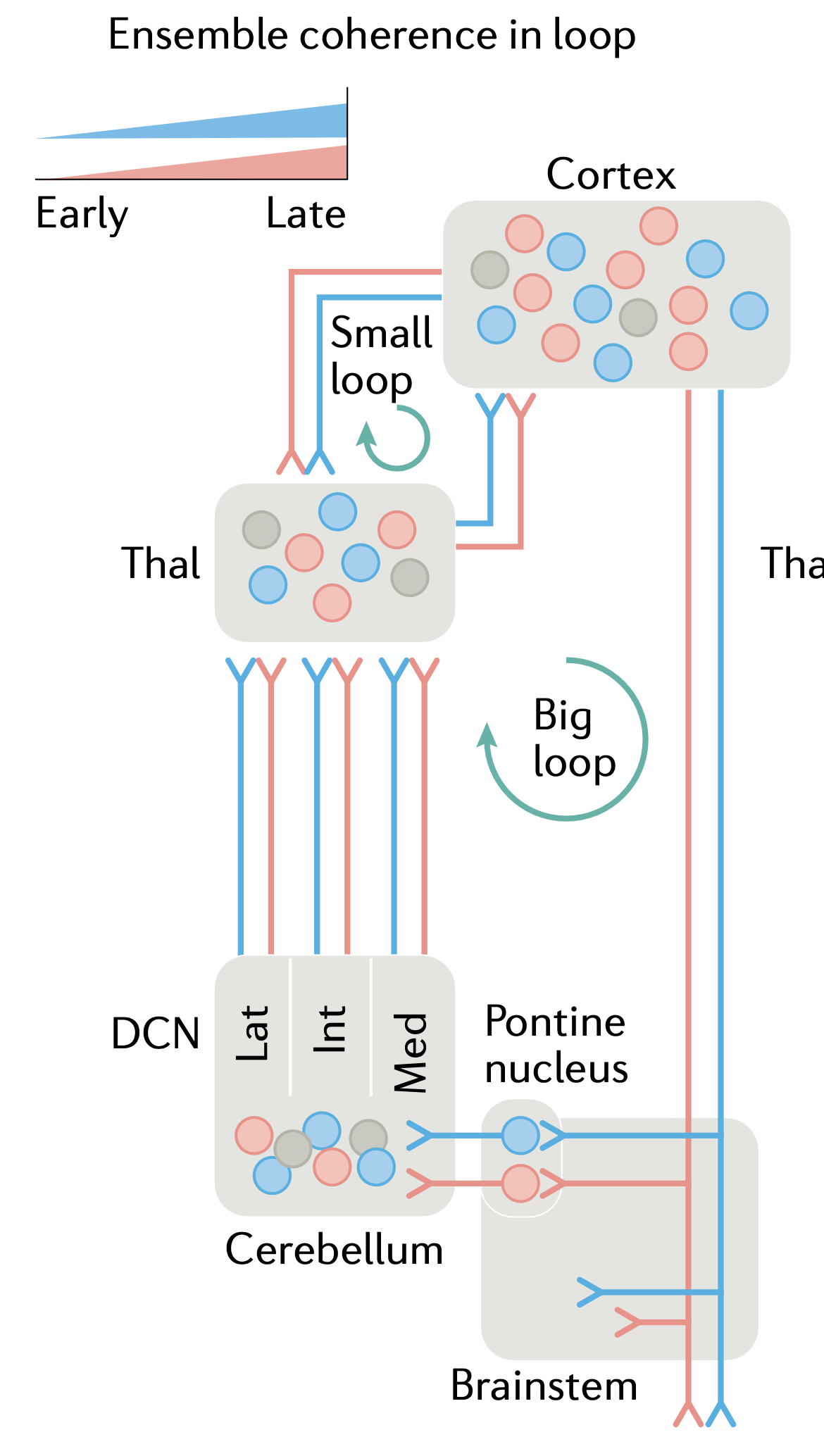
前頭葉皮質が下行運動コマンドを駆動する時刻と、これらの上行経路からの遠心性コピー信号が実際に生成された実際の低レベル運動コマンドを反映する時刻との間には、自然な遅れが存在します(上記のすべての領域が大きく寄与し、形成されています)。この遅延は、予測学習を推進するために必要な [[temporal derivative]] ダイナミクスを提供します。この場合、運動皮質は、送信されたコマンドに応答して運動システムが何を行うかを予測するために効果的に学習します。この仮説は、[[#figure_big-loop]] ([[@ArberCosta22]] より) に示されている下降および上昇モーター信号の「大きなループ」と一致します。
{id=”figure_big-loop” style=”高さ:30em”}
さらに、体性感覚皮質 (S1) から運動皮質 (M1) までの重要なドライバー様経路があり、皮質皮質経路を直接経由する経路 ([[@PetrofViaeneSherman15]]) と、視床後内側視床 (POm) を経由する視床経路を経由する経路 ([[@MoSherman19]]) の両方があります。 S1 が、遠心性コピー信号や、脊髄と脳幹の運動相乗効果に対するフィードバック制御の提供に関与するすべての固有受容信号を含む、運動動作の感覚的結果を反映する広範な進行中の入力も受け取ることを考えると、これは、皮質がそのような信号をオンラインフィードバック制御自体に利用することと、VL ベースのビッグループ経路で得られるのと同じ種類の予測学習メカニズムを介して、運動表現の学習を促進するためにこれらの信号を使用することの両方を行う機会を提供します。
したがって、予測学習を介して新皮質における運動学習を訓練する際のこれらの体性感覚信号の役割は、感覚結果信号の予測学習を介して小脳が学習する方法と同形である。小脳は、進行中の運動動作をきめ細かいタイムスケールでオンライン調整できるように組織されていますが、新皮質は一般に、より遅いタイムスケールで動作し、[[bidirectional connectivity]] および [[constraint satisfaction]] 処理を使用して動作を選択し形成することがより重要です。
| [[#figure_big-loop]] の「小さなループ」には、さまざまな視床ニューロンと領域、特に腹側前部および内側 (VA、VM) が関与しており、これらはマトリックス状であり、運動皮質の最も表層に広く拡散した接続を送ります。したがって、これらの領域は運転学習には適していません(興奮性のドライバー入力が得られません)が、活動の準備状態から実際に運動活動を駆動する活動パターンへの移行を促進する上で重要な役割を果たす可能性があります([[@EconomoViswanathanTasicEtAl18]]; [[@GuoYamawakiSvobodaEtAl18]]; 詳細については、前頭前野ページの [[prefrontal cortex | systems-level dynamics]] を参照)。 |
運動活動のより抽象的な状態空間分析では、準備運動活動が、実際に下位レベルの運動出力経路を駆動する活動状態と比較して「空白空間」に存在することが一貫して示されています ([[@ChurchlandShenoy24]])。これらの出力経路は、5b 層の PT (錐体路) ニューロン (特に下層 5b; [[@EconomoViswanathanTasicEtAl18]]) によって駆動され、脱抑制性大脳基底核入力を受け取る VA 視床領域から広く突き出たマトリックス状経路によって (頂端房内で) 選択的に標的とされます ([[thalamus#frontal thalamus]] を参照)。したがって、BG 入力によるこれらの PT ニューロンの排他的な活動ほど単純ではありませんが、準備ヌル空間からモーター出力空間への移行は、これらの BG 信号によって促進される可能性があります。
最終的な運動出力パラメータに関連した運動皮質表現の性質については、多くの研究が行われています。あらゆる種類の関連信号が個々のニューロンからデコードされていますが、いつものように、全体的な [[distributed representations]] の方が関連性が高くなります ([[@GeorgopoulosCarpenter15]])。 [[@^AflaloGraziano06a]] は、最終的な手の姿勢が変動の大部分を占めることを示し、これは上で議論した筋肉の相乗効果の一般的な性質と一致しています。ただし、速度、空間の曲率、距離、力などの他の要素もコード化されました。
別の研究では、より複雑な一連の運動動作が必要な場合を調査し、姿勢構成の時間的シーケンスが指定されることを示しました(つまり、時空間相乗効果; [[@OverduindAvellaRohEtAl15]])。
より抽象的なレベルの分析では、[[@^MeyerSmithWright82]] は、人間の運動の速度と精度に関する広範な精神物理学的文献を総合して、前半の初期加速段階とそれに続く後半の対称的な減速段階を持つ曲線として力と持続時間パラメーターを指定する対称インパルス制御モデルを開発しました。これらの曲線の力と時間のパラメータはどちらも人間が制御できます。弾道の動きが約 260 ミリ秒未満で行われ、時間の許す限りその後に視覚的に修正された動きの更新が複数回繰り返されるという証拠があります ([[@MeyerSmithKornblumEtAl90]] も参照)。