compcogneuro/web: neuron-detector
このページは外部資料の日本語訳です。原文の見出し順と本文順を保ち、コード・URL・出典表記はできるだけ原形のまま残しています。
出典とライセンス
原典: https://github.com/compcogneuro/web/blob/main/content/neuron-detector.md
ライセンス: Text: CC BY 4.0; code: BSD 3-Clause。このページは日本語翻訳であり、変更点は翻訳とサイト内整形です。
+++ Categories = [“Activation”, “Neuroscience”] bibfile = “ccnlab.json” +++ 検出のプロセスの観点から [[neuron|neural]] 関数を理解するための、高レベルで概念的な便利な方法があります。ニューロンは他のニューロンから何千もの異なる入力信号を受け取り、自分にとって「意味のある」特定のパターンを探します。非常に簡単な例としては、空気をサンプリングして明らかな煙の痕跡を探す煙探知機があります。これらが指定されたしきい値制限を超えると、アラームが鳴ります。同様に、ニューロンには閾値があり、この閾値を超えるほど重大な何かを検出した場合にのみ、他のニューロンに「警報」信号を送信します。アラームは 活動電位 または スパイク と呼ばれ、ニューロン間の通信の基本単位です。
{id=”figure_detector” style=”高さ:20em”}
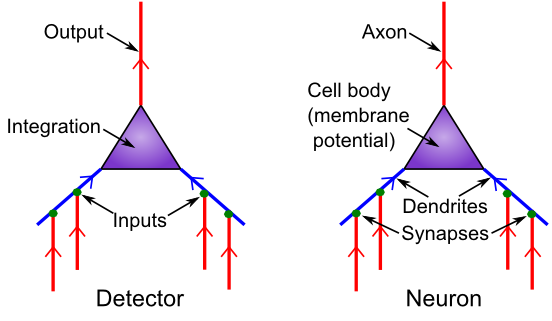
[[#figure_detector]] は、神経生物学とそれらが提供する検出機能との対応を示します。 シナプスは、送信ニューロン (アラームを発して信号を送信するニューロン) と 受信ニューロン (その信号を受信するニューロン) の間の接続ポイントです。ほとんどのシナプスは、大きな枝分かれした木である 樹状突起 上にあり (「樹状突起」という言葉は、木を意味するギリシャ語の「デンドロス」に由来します)、ニューロンはそこですべての入力信号を統合します。
主要な川に流入する支流と同様に、これらの信号はすべて樹状突起の幹に流れ込み、細胞体に流れ込み、そこで信号の最終的な統合が行われます。しきい値処理は、軸索と呼ばれるニューロンの出力端の最初の部分で行われます (この開始場所は 軸索丘 と呼ばれます – どうやら小さな丘か何かのように見えます)。また、軸索は広く分岐し、他のニューロンの樹状突起上にシナプスの反対側を形成し、次の伝達連鎖を完成させます。そしてさらに進みます。
最も基本的なレベルでは、ニューロンの機能生物学について知っておくべきことはこれだけです。ニューロンは入力を受け取り、それらを統合し、統合された入力が出力信号をトリガーするのに十分な強さであるかどうかを判断します。ただし、これらの入力信号の性質に関しては、統合プロセスが全体的な入力信号強度の大きな変化にうまく対処できるようにするなど、神経機能に影響を与える追加の生物学的特性がいくつかあります。
ニューロンへの入力信号の主なソースは 3 つあります。
-
興奮性入力: これらは、他のニューロンからの「通常の」最も一般的なタイプの入力 (全入力の約 85%) であり、受信ニューロンを興奮させる効果があります (閾値を超えて「警報」を発する可能性が高くなります)。それらは、神経伝達物質グルタミン酸によって開かれるAMPAと呼ばれるシナプスチャネルを介して伝達されます。
-
抑制性入力: これらは入力の残りの 15% であり、興奮性入力とは逆の効果を持ちます。これらはニューロンが発火する可能性を *低くし、興奮を抑制することで統合プロセスをより堅牢にする役割を果たします。脳には、この抑制性入力を生成する 抑制性介在ニューロン と呼ばれる特殊なニューロンがあります (詳細については、[[inhibition]] を参照)。この入力は、神経伝達物質 GABA によって駆動される GABA シナプス チャネルを介して入ります。
-
リーク入力: これらは常に存在してアクティブであるため、厳密には入力ではありませんが、興奮を打ち消してニューロン全体のバランスを保つことにより、抑制性入力と同様の機能を果たします。生物学的には、リーク チャネルは カリウム チャネル (K) です。
抑制性と興奮性の入力は、皮質の「異なる」ニューロンから来ます。特定のニューロンは、興奮性または抑制性の出力のいずれかのみを他のニューロンに送信でき、両方には送信できません(他の脳領域のニューロンはこの制約に違反しますが、新皮質錐体ニューロンはそれに従うようです)。この制約がもたらすさまざまな影響を本文全体で見ていきます。
最後に、正味シナプス効率 または 重み の概念を導入します。これは、送信ニューロン活動信号がシナプス接続を介して受信ニューロンに与える可能性のある合計の影響を表します。 シナプスの重みは、計算認知神経科学の全分野において最も重要な概念の 1 つです!。これは、[[abstract neural network]] モデルの重要な要素でもあります。
生物学的には、この重みは、神経伝達物質を放出する送信ニューロンの活動電位の正味の能力と、その神経伝達物質がシナプス後側のシナプスチャネルを開く能力(開くことができるそのようなチャネルの総数を含む)を表します。興奮性入力の場合、重みは送信ニューロンによってシナプスに放出されるグルタミン酸の量と、シナプスの受信ニューロン側の AMPA チャネルの数と有効性に依存します ([[neuron#figure_synapse]])。
計算上、重みによってニューロンが何を検出しているかが決まります。強い重み値は、ニューロンがその特定の入力信号に対して非常に敏感であることを示し、一方、低い重み値は、その入力が比較的重要ではないことを意味します。学習とは基本的に、送信ニューロンと受信ニューロンの神経活動パターンに応じてこれらのシナプスの重みを変更するだけです。
つまり、あなたが知っているすべてのもの、脳内のすべての大切な記憶は、シナプス重みのパターンとしてエンコードされています! この検出プロセスは、[[categorization]] をサポートしており、また、線形代数用語で、特定の dimensions、bases、または axes (目的にとってはすべて同義語) に沿った高次元入力空間の projection もサポートしています。したがって、この検出プロセスはまさにニューラル計算の基本エンジンであり、さまざまな方法で説明できますが、すべてここで分解する同じ本質的なプロセスに相当します。
リンク
[[Intro Book]] の次: [[Neuron]]