compcogneuro/web: prefrontal-cortex
このページは外部資料の日本語訳です。原文の見出し順と本文順を保ち、コード・URL・出典表記はできるだけ原形のまま残しています。
出典とライセンス
原典: https://github.com/compcogneuro/web/blob/main/content/prefrontal-cortex.md
ライセンス: Text: CC BY 4.0; code: BSD 3-Clause。このページは日本語翻訳であり、変更点は翻訳とサイト内整形です。
+++ Categories = [“Rubicon”, “Neuroscience”] bibfile = “ccnlab.json” +++ 前頭前皮質 (PFC) は、生物の全体的な行動の多くを導く上で、[[basal ganglia]] (BG) との主な共謀者です。それは 前頭 [[neocortex]] の前部である [[anatomy|anatomically]] です。後部前頭皮質には、脊髄まで突き出て運動動作を直接駆動できるニューロンを備えた一次運動野と二次運動野があります。 PFC は、後部皮質領域への広範な接続とともに、これらの運動前頭領域に直接「トップダウン」投影を提供し、行動と認知を「より高いレベル」で導きます。
| PFC のこの役割は、会社の役員 ([[@MillerCohen01]]) に例えて、執行機能 と呼ばれます。しかし、これには、この PFC 幹部の役割において、無傷の人間の脳の説明されていない認知能力が完全に呼び出されることで、無限後退する危険性があります。脳の自律的な計算モデルを作成する場合、PFC ([[@HazyFrankOReilly06]]) に住むこの「ホムンクルス」 (「小さな人」) から逃れることはできません。したがって、我々の課題は、程度の差こそあれ他の霊長類や他の哺乳類とほぼ同様の生物学的システムから、ヒトに特有のオープンエンド型 [[artificial intelligence | general intelligence]] がどのようにして現れるのかを理解することです。 |
[[Rubicon]] フレームワークでこの課題に対して採用されるアプローチは、まず、考えられるさまざまな行動方針の中から選択し、選択された目的を達成するために行動と認知を導く能力を PFC がどのようにサポートするかを理解することです。この目標主導型の機能は、大脳基底核および他の皮質下領域の相互接続された領域と相乗的に機能する、PFC領域の分散ネットワークによってサポートされています。
{id=”figure_odr-delay” style=”高さ:40em”}
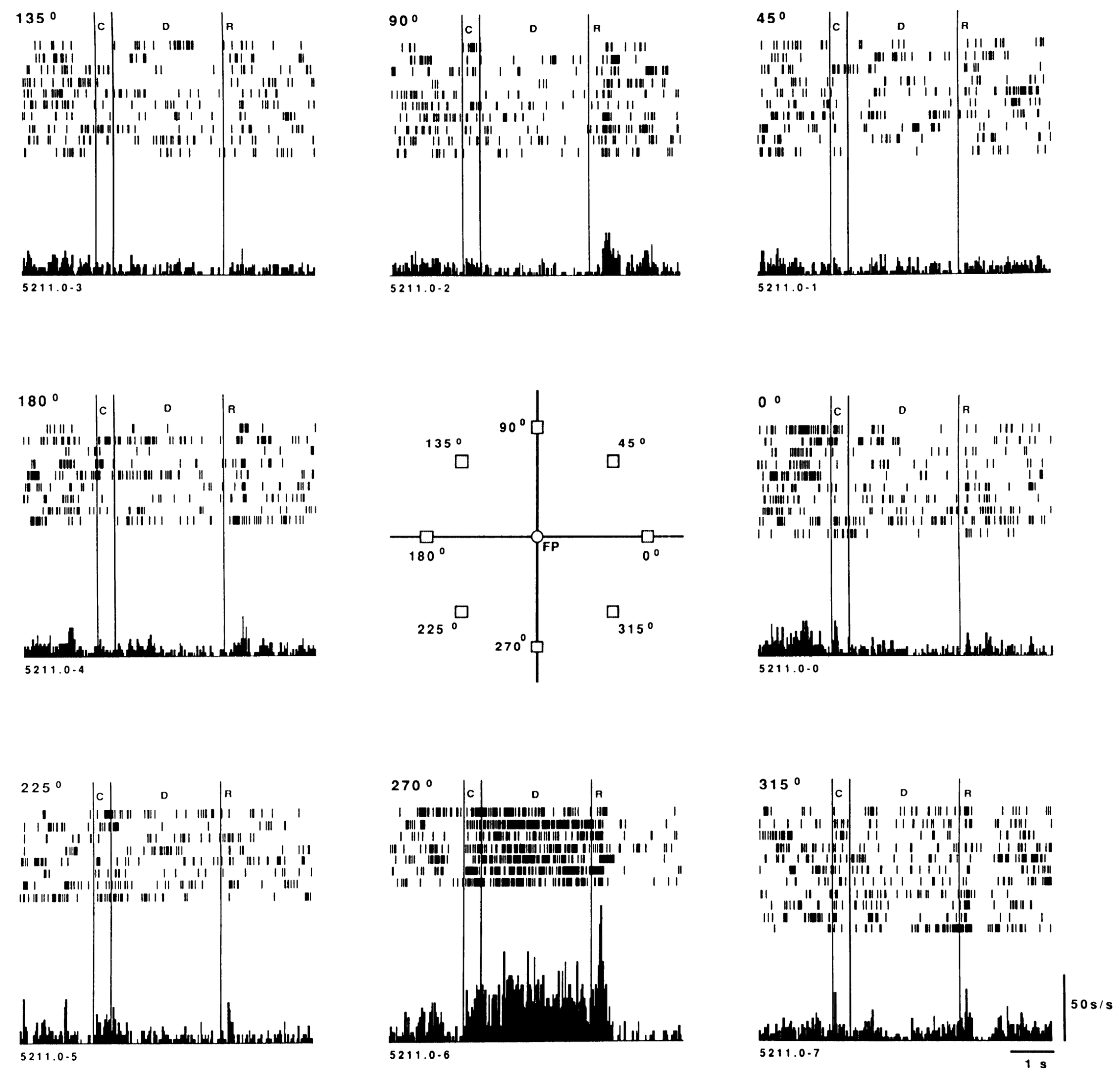
PFC がもたらす独自の「スーパーパワー」は、長期にわたって堅牢な方法で神経発火を維持する能力 (つまり、作業記憶; [[@BaddeleyHitch74]]; [[@MiyakeShah99]]; [[@KubotaNiki71]]; [[@FusterAlexander71]]; [[@Goldman-Rakic95a]]) であり、これは注意を集中させ続ける「接着剤」です。選んだゴール。 [[@^FunahashiBruceGoldman-Rakic89]] の典型的な例については、[[#figure_odr-delay]] を参照してください (馴染みのない場合は、[[experimental-methods#peristimulus time histogram (PSTH)]] でプロットを説明します)。この能力の神経基盤は、高レベルの [[neuron channels#NMDA]] チャネルを備えた強力な双方向回路に由来しており、高度な [[stable activation]] ([[@LismanFellousWang99]]; [[@BrunelWang01]]; [[@SandersBerendsMajorEtAl13]])。 [[dopamine]] や [[acetylcholine]] などのさまざまな神経調節物質も重要な役割を果たします ([[@CoolsArnsten22]])。
この点で PFC が何をしているかを理解するための最も顕著な方法は、PFC が睡眠中に非活性化されるという事実 ([[@HobsonPace-Schott02]]) から得られます。つまり、夢を見ている間は一貫した集中力を維持できないということは、PFC の重要な貢献を示しています。特に後新皮質は、意味的な関連性や潜在的なつながりや洞察を生成することに優れていますが、すぐに気が散ってしまい、目の前のタスクに集中し続けるための PFC がなければ意識の流れの軌道をたどります。
通常、実験研究では遅延期間を使用します。この期間では、PFC が確実に動作するように、常に変化するタスク情報を少なくとも 1 秒以上維持する必要があります ([[#figure_odr-delay]] など)。それ以外の場合、BG ページでレビューされているように、[[basal ganglia]] は通常、前頭葉の助けなしで管理できます。しかし、動物の行動の多くを決定する比較的単純で再現可能な手順が必要なこのような研究には限界があるため、PFC や目標主導の内生的に制御される行動の本質のほとんどが欠けている可能性があります。幸いなことに、計算モデルではこれらの制限を回避して、より自由な動作を理解することができます。
以下のセクションでは、多くの異なる領域にわたる PFC の機能構成の詳細な考察から始まり、その後、[[thalamus]] の重要な役割と大脳基底核によるその調節も関与する、堅牢な能動的維持をサポートする神経機構の概要を説明します。
PFC の機能組織
{id=”figure_goal-bio” style=”高さ:20em”}
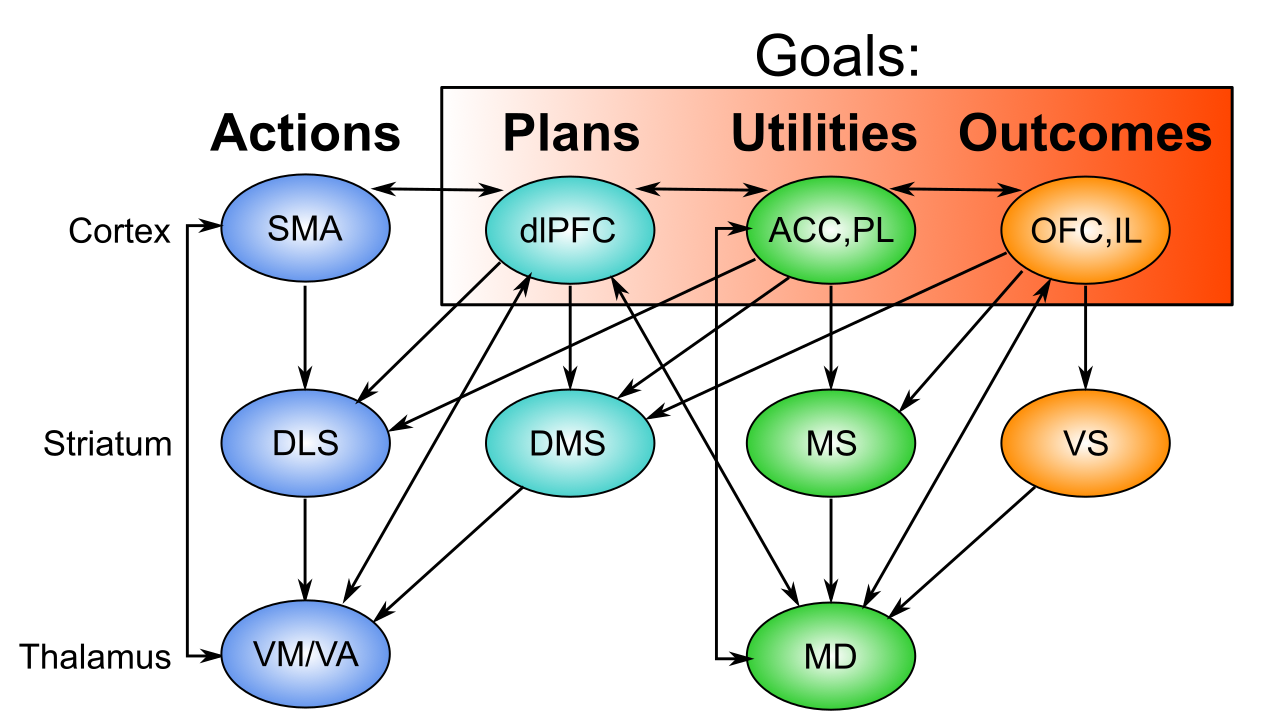
[[Rubicon]] 目標駆動フレームワークは、[[#figure_goal-bio]] に示すように、行動目標の分散表現を構成する PFC 領域の 3 つの異なるグループを区別します。
-
OFC (眼窩前頭皮質) および IL (辺縁下皮質) は、さまざまな抽象化レベルでの動機付けに顕著な結果を表す腹側および内側 PFC 領域であり、標準的な条件付け用語では US (無条件刺激) と呼ばれます。これらは、目標主導の行動に取り組むときに得ようとしているものです: 食べ物、水、賞賛などです ([[emotion]] を参照)。
-
ACC (前帯状皮質) および PL (辺縁前皮質) は、目標の達成に関連する_コスト_ (ACC における努力、リスク、不確実性)、および目標の達成に関連する純利益とコストの_ユーティリティ_ (PL における) を表す、背内側 (ACC) および前内側 (PL) の PFC 領域です。したがって、PL は、目標主導型の行動にとって最も重要な統合領域であり、特定の目標を実際に達成することの正味推定値を提供します。
-
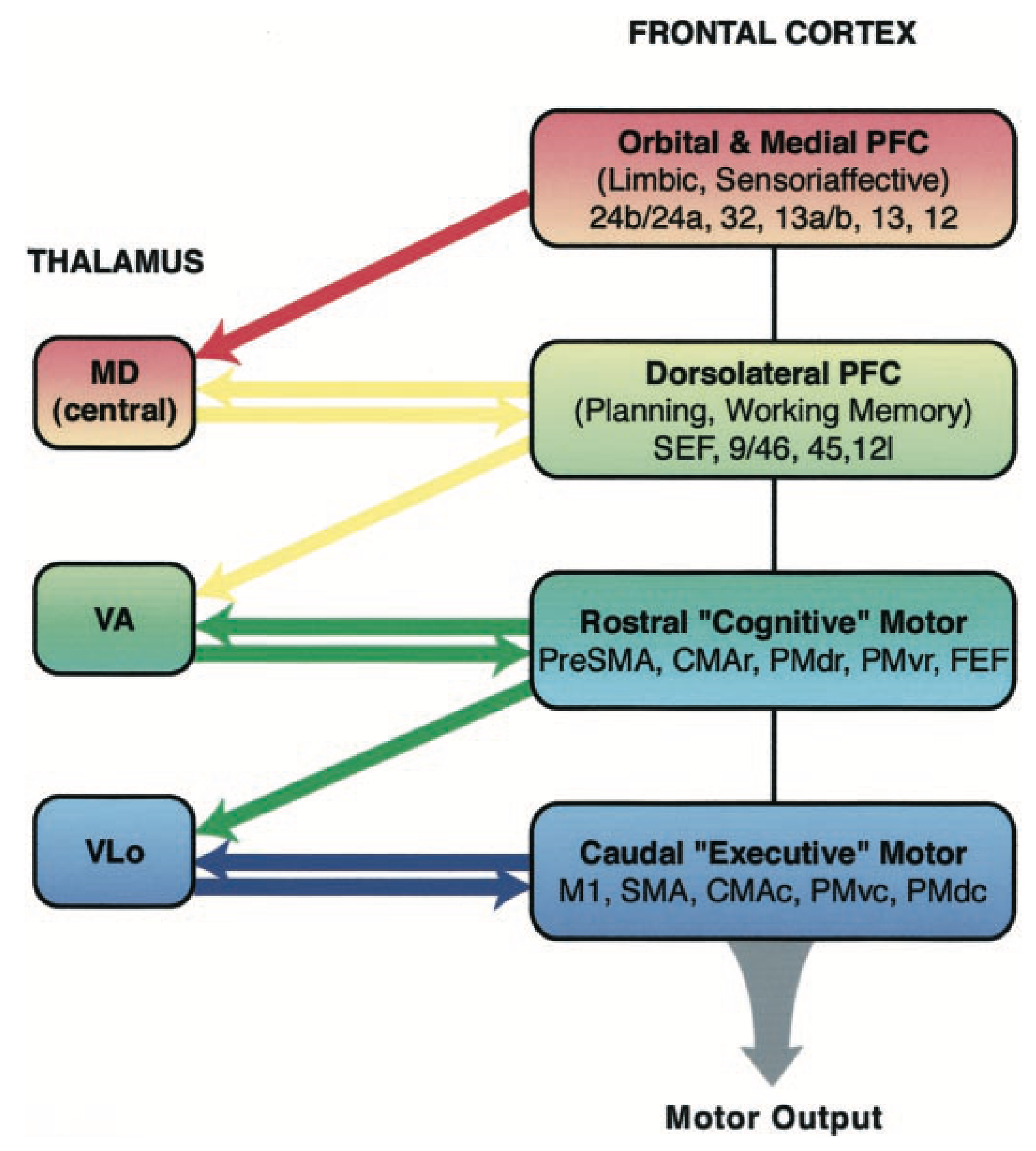
dlPFC (背外側 PFC) (げっ歯類の ALM (前側方運動)) は、望ましい結果を得るために効果的である可能性のある高レベルの行動計画を表します。これは、これらの計画に関連するコストなどの見積もりを推進するために ACC に広範囲に投影し、目標が選択されて実行されると、この広い概念では運動前野を含む SMA (補足運動野) の行動表現にトップダウンのバイアスを提供します。これらの高次の運動野は同様に、一次運動野における個々の運動表現を駆動します。
したがって、これらのさまざまな PFC 領域にわたって、目標の評価と実行に必要な重要な情報がすべてエンコードされます。堅牢なアクティブ メンテナンス (ワーキング メモリ) 機能により、これらの領域全体に分散された目標表現が時間の経過とともに持続し、目標を達成するための適切な動作が推進されます。
また、[[#figure_goal-bio]] に示されているように、各 PFC 領域は、[[basal ganglia]] (BG) の対応する領域および視床のさまざまな領域と広範囲に相互接続されており、複数の平行ループ ([[@AlexanderDeLongStrick86]]; [[@FosterBarryKorobkovaEtAl21]]) を形成します。また、腹側 - 内側 OFC、IL、PL、および図 ([[@HaberFudgeMcFarland00]]) に示すように、ACC 野は、dlPFC および運動野と相互作用する領域に非対称に投影されますが、その逆はありません。
| これらのBGと視床結合の機能的役割はかなりの議論の的となっているが、最近の神経科学データ([[#systems-level-dynamics | below]]でレビュー)は、BGがPFCにおける持続的な神経発火をゲートまたはトグルできるという考えを裏付ける、初期の計算上のアイデアと収束しつつある。ただし、このゲート機能は主に、上記の腹側/内側 PFC 野全体で行われる最初の目標選択プロセスに関連していますが、前頭運動野と相互接続する背外側 BG 野は、BG での学習に基づいて皮質での学習を促進できる調節信号を提供すると考えられます。 |
{id=”figure_pfc-comp” style=”高さ:40em”}
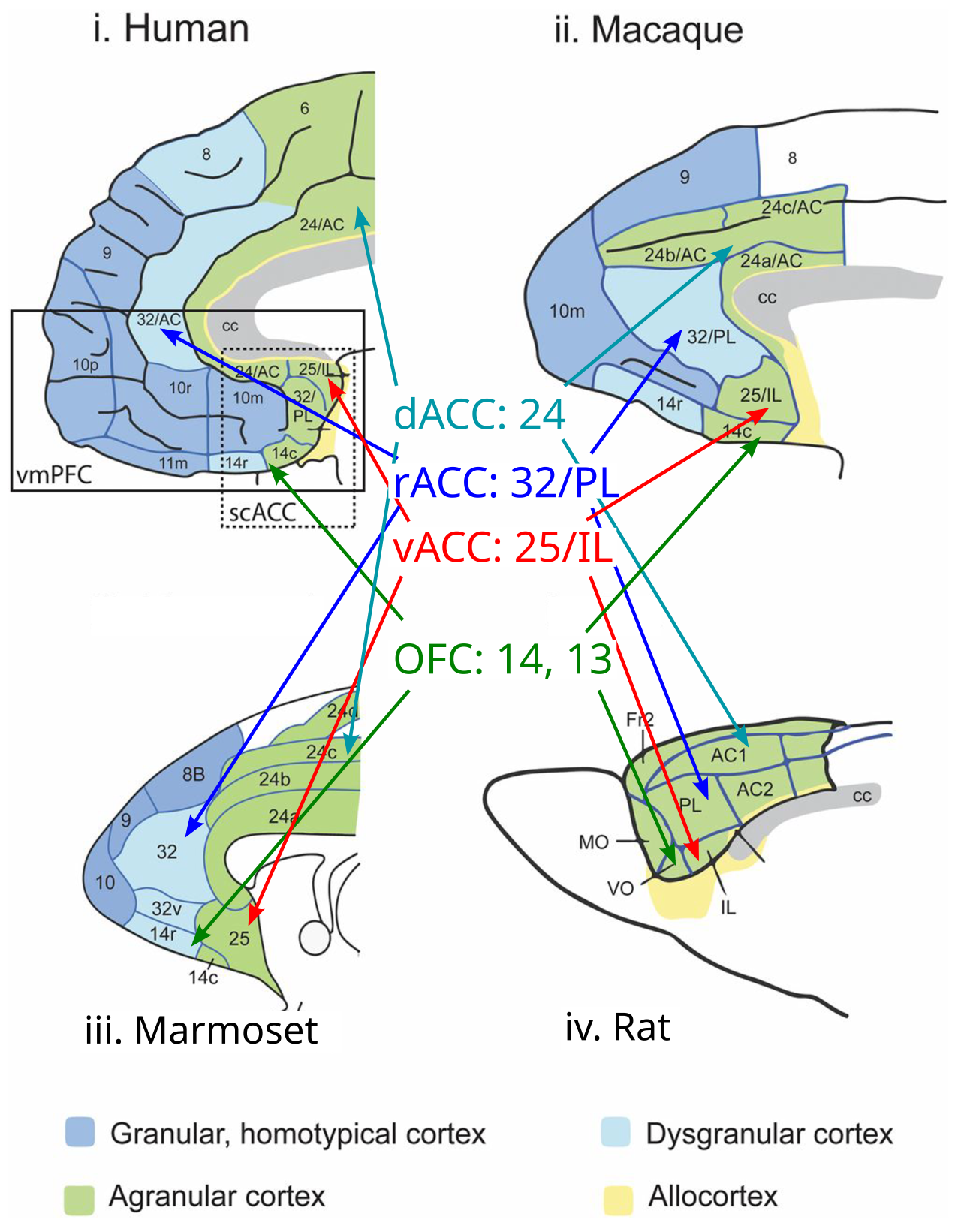
| [[#figure_pfc-comp]] は、ラットからヒトまでのさまざまな種 ([[@RobertsClarke19]]) にわたる中核となる目標主導型 PFC 領域を示します。途中に 2 つの「中間」種 ([[evolution | evolutionarily]] より古い霊長類であるマーモセットと、ヒトを含む類人猿の系統に近いマカク) が含まれます。目標主導型 PFC 野はラット ([[#figure_pfc-rat]]) の PFC の大部分を構成しますが、霊長類には追加の背側 / 側方 PFC 野があり、より精緻な行動計画をサポートします。 |
{id=”figure_pfc-rat” style=”高さ:20em”}
この進化の軌跡から、目標主導型 PFC 領域は、進化の過程を通じて保存されている PFC 機能の最も重要な「中核」の側面を実際にサポートしていると推測できます。これは、げっ歯類の PFC に計算モデリングの取り組みを集中させることで、PFC 機能の最も重要な側面を捉えることができ、その後のモデルで霊長類の脳の精緻な計画と運動制御能力を捉えることができることを意味します。
{id=”figure_pfc-fun” style=”高さ:40em”}
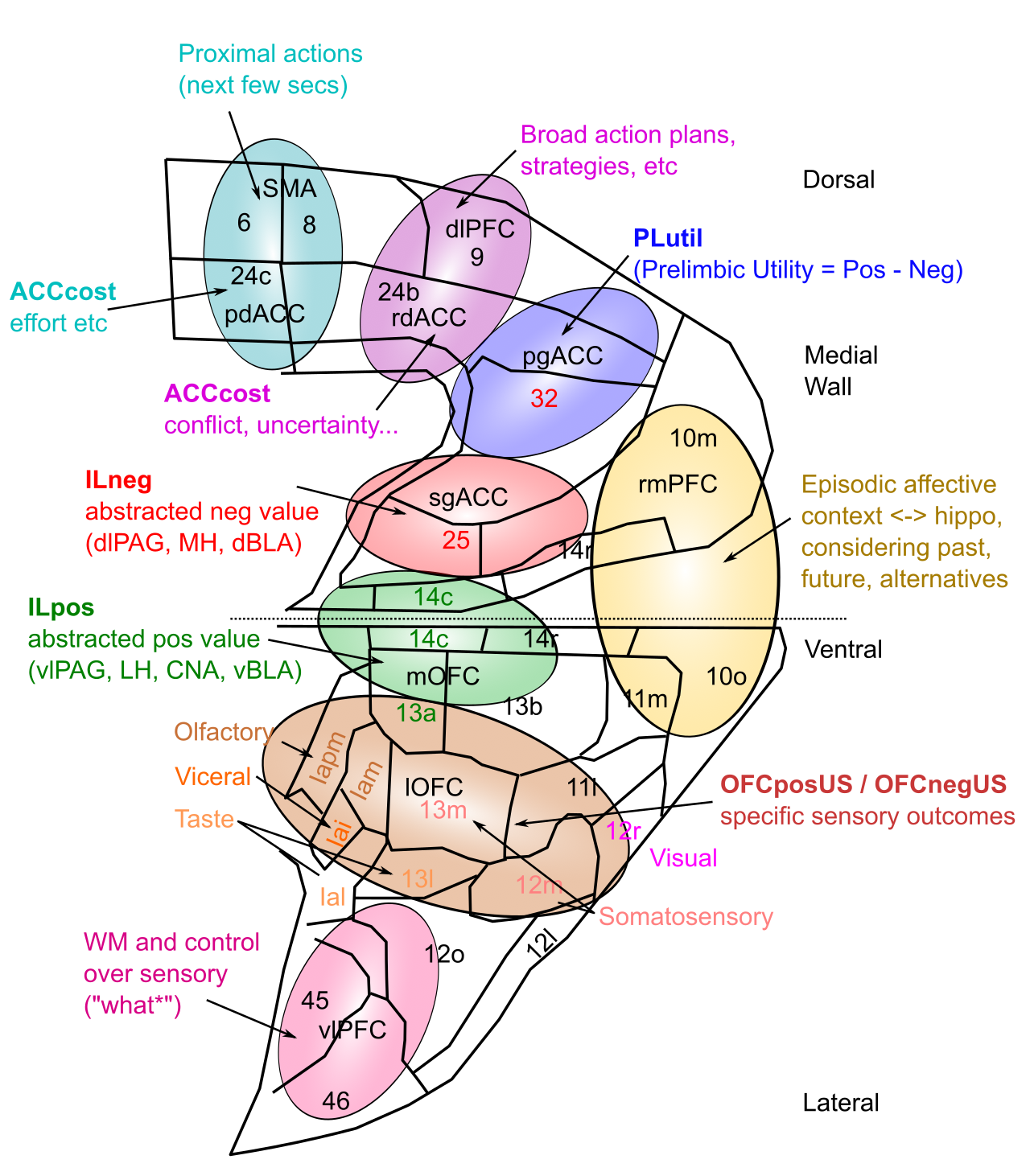
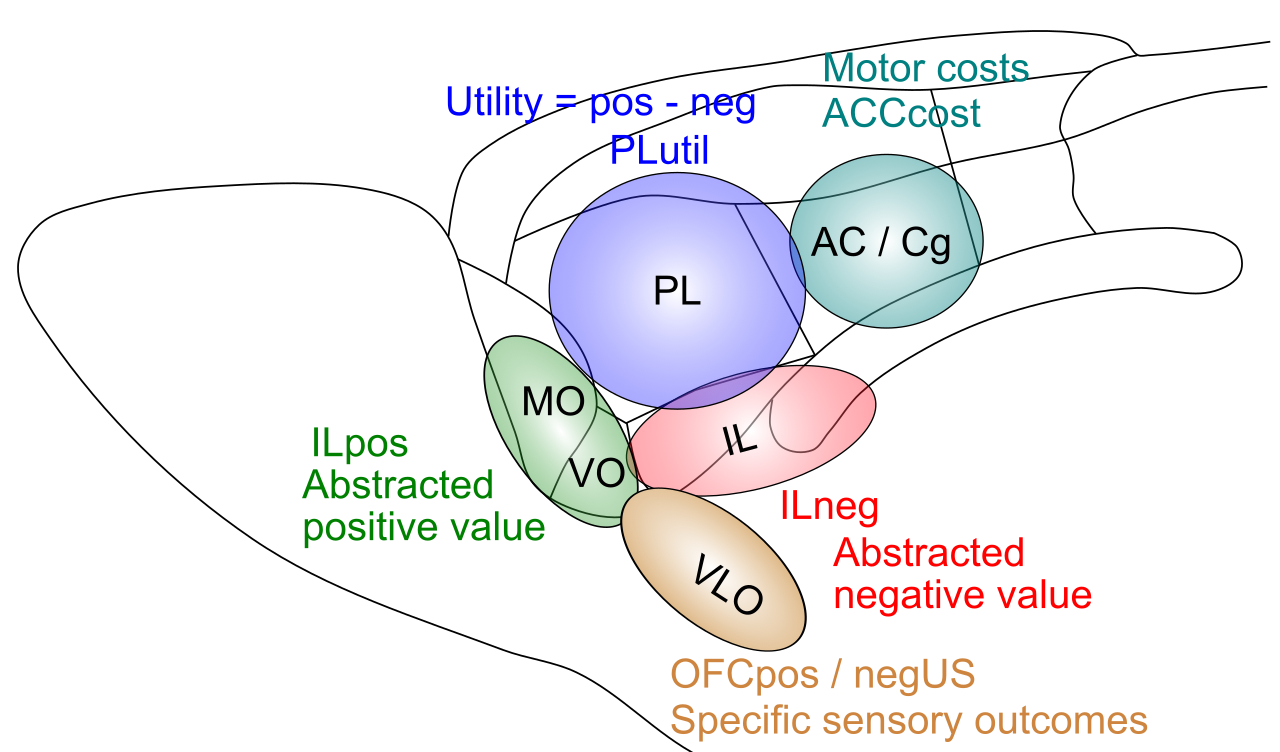
[[#figure_pfc-fun]] は、霊長類 PFC の精緻なバージョンを提供し、[[@^OngurPrice00]] で説明される皮質下領域との接続に基づいて、より詳細な機能ラベルを割り当てます。 [[emotion]] で説明されているように、これらの皮質下領域は、強力な進化制御下にある、比較的明確に限定された神経反応特性を持ち、対応する PFC 領域に学習を定着させて、生存関連因子の制御に必要な関連情報を表すことができます。
ニューラルコーディングデータ
腹側および内側 PFC 野の上記の機能解剖学と一般的に一致する、複数の種にわたる豊富な神経記録データがあります。しかし、動物のデータの多くは個々のニューロンの記録からのものであり、必然的に、これらの異なる PFC 領域すべてにわたるニューロンが、上記のフレームワークの特定の領域に帰せられる機能変数(たとえば、[[@KennerleyBehrensWallis11]] およびその中の参考文献)に対してある程度の感受性を示すことが示されています。
[[distributed representations]] で説明されているように、個々のニューラル コーディングのこの「混合選択性」は遍在しています。これは、特定の脳領域の独特のコーディング特性を理解するのに _表現類似性分析 (RSA) が最適なツールである理由も説明しています。
{id=”figure_hunt-rsa” style=”高さ:40em”}
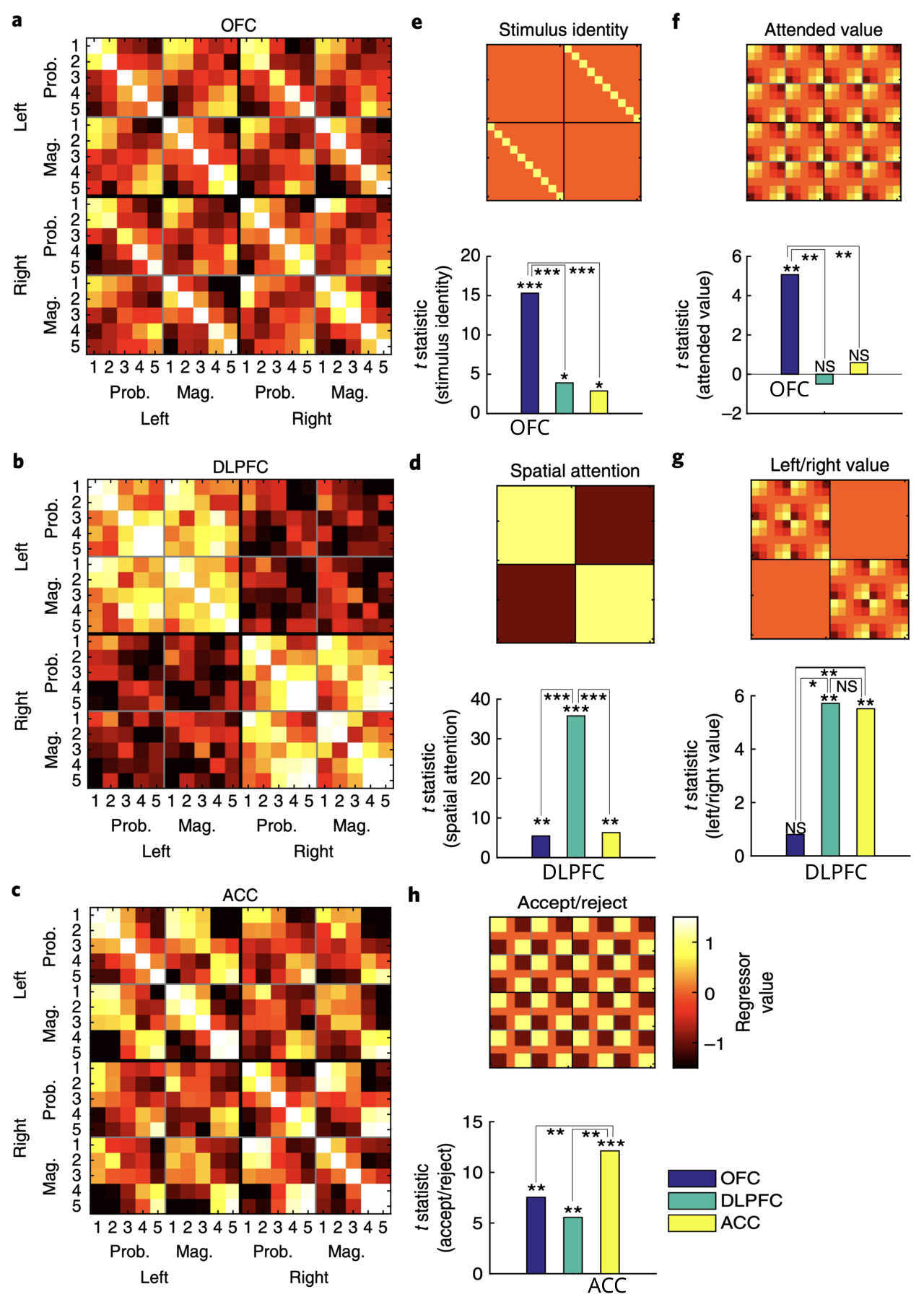
| 幸いなことに、[[@^HuntMalalasekeradeBerkerEtAl18]] は、意思決定タスクで dlPFC、ACC、および OFC 領域の [[experimental methods#Representational similarity analysis (RSA) | RSA]] 分析を実行し、[[#figure_hunt-rsa]] に示されているように、これらの領域の機能特化に対する顕著なサポートが見つかりました。具体的には、OFCニューロンは、結果の価値を表すことと一致して、サルが現在参加している刺激の刺激の同一性と報酬値の両方をコード化した。代わりに、DLPFC ニューロンは、実行する必要がある左 / 右の選択アクション (実行前、つまりアクション プラン)、および ACC とともに、これらのアクションの選択の全体的な値をエンコードしました。最後に、ACC は、すべての変数を統合した最終的な承認/拒否の決定を表します。 |
これらの領域間で [[bidirectional connectivity]] を介して動作する並列 [[constraint satisfaction]] プロセスと一致して、これらの神経反応は領域全体で並行して出現しました。したがって、各領域は全体的な目標の特定の側面をコード化するために特化されていますが、すべてが連携して、それぞれの制約を満たす計画に収束します。
サルが異なる選択肢を検討したとき、[[@^RichWallis16]] による OFC でも同様の動態が観察され、これは任意の時点で単一の選択肢を検討するという要件と一致しています。これは、制約満足プロセス ([[@HerdKruegerNairEtAl21]]) で [[binding problems]] を回避するために必要です。
アクティブメンテナンスのメカニズム
{id=”figure_pfc-maint-lamina” style=”高さ:40em”}
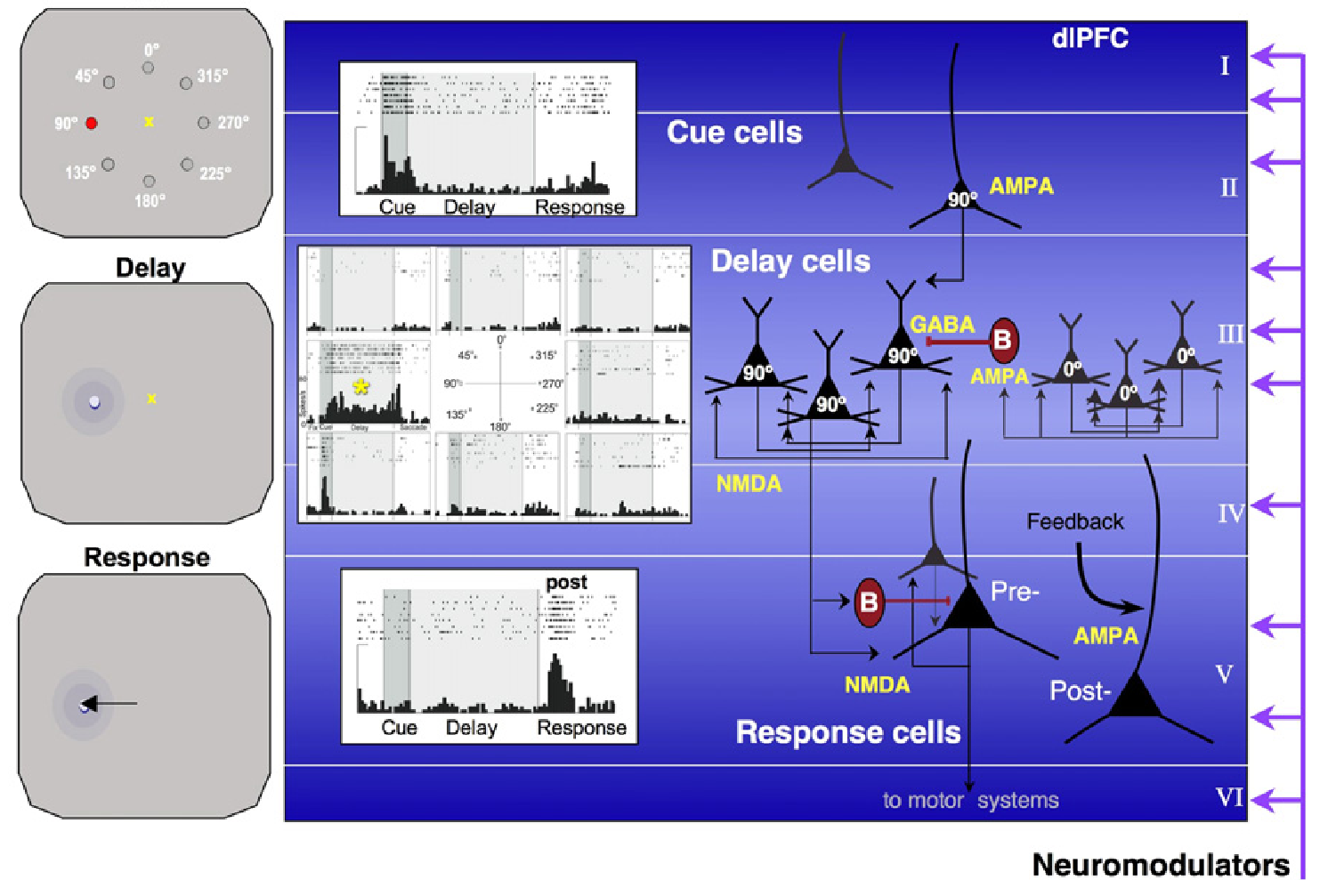
[[@^ArnstenWangPaspalas12]] の [[#figure_pfc-maint-lamina]] は、双方向の興奮性接続が時間の経過とともにどのように神経活動を堅牢に維持するかについて広く受け入れられているフレームワークを示しています。この場合、標準的な眼球運動遅延応答タスク内で合図位置をエンコードしています。具体的には、PFC内の深層3錐体ニューロンの特殊な集団が同定されており、これは広範な側方相互興奮性(反復性)結合性と高濃度のNMDA受容体を持っています([[@KritzerGoldman-Rakic95]]; [[@WangMarkramGoodmanEtAl06]])。
この接続パターンは霊長類で顕著な進化的拡大を遂げており ([[@Elston03]]; [[@WangYangWangEtAl13]])、いくつかの研究では、NMDA 受容体遮断が複数の種で作業記憶のパフォーマンスを損なうことが示されています ([[@KrystalAbi-SaabPerryEtAl05]]; [[@MoghaddamAdams98]]; [[@RobertsShafferSeymourEtAl10]]; [[@WangYangWangEtAl13]])。げっ歯類では、次のセクションで検討した証拠は、能動的な維持が第 5 層の PT ニューロン (これも NMDA で強化された反復接続; [[@WangStradtmanWangEtAl08]]) と視床を通る双方向興奮性ループに依存しているのに対し、霊長類は深部の第 3 層ループに基づいて神経発火をより堅牢に維持できる可能性があることを示しています。
[[#figure_pfc-maint-lamina]] に示される層の特化は、[[neocortex]] 接続の標準パターンと一致しており、入力が直接および第 4 層を介して間接的に表層を活性化します。PFC からの皮質下の出力は深層から発生し、大きな層 5b の出力ニューロンが直接運動レベルの出力を提供します (つまり、それらの軸索は脊髄への錐体路 (PT) の投影を構成します)。これらの第 5 層ニューロンはまた、大脳基底核および他の皮質下の標的にも投射し、次に概説するように視床皮質ループを形成します。
出力応答を駆動することに加えて、層 5b 出力ニューロンは、識別された層 5b ニューロン ([[@SommerWurtz00]]) におけるこれらすべての発火パターンの明確な記録によって明らかになったように、感覚入力信号と持続的な能動維持信号の両方も送信します。これは、第 2 層および第 3 層のニューロンから第 5b 層への投影の異なるパターンから生じる可能性があり、PFC 活動のあらゆる側面を皮質下システムで利用できるようにするのに計算的に役立ちます。
システムレベルのダイナミクス
PFC 内の活動のダイナミクスは、[[thalamus]] からの広範な投影による強い影響を受け、さらに [[basal ganglia]] や [[cerebellum]] などの他の脳領域からの入力や、他の神経調節入力の影響を受けます。 PFC は、これらの分野やその他の分野に広範な予測を送ります。ここでは、これらのシステムレベルの相互作用に関するデータを確認します。
まず、視床と PFC の間の広範な双方向興奮性接続により、これらの視床皮質ループが能動的な維持、および PFC で情報が更新されるときと維持されるときの動的な制御において重要な役割を果たす計算モデルの開発に多くの人が導きました ([[@DomineyArbib92]]; [[@DomineyArbibJoseph95]]; [[@BeiserHouk98]]; [[@FrankLoughryOReilly01]]; [[@OReillyFrank06]]; [[@AshbyEllValentinEtAl05]])。
たとえば、[[PBWM]] モデル (前頭前皮質大脳基底核の作業記憶) [[@OReillyFrank06]]) は、広く使用されている [[LSTM]] モデル (長短期記憶、[[@HochreiterSchmidhuber97]]) からの計算上の洞察と、これらの視床皮質回路および大脳基底核を介した脱抑制制御との間の直接的な関係を引き出しています。
LSTM モデルは、情報が PFC で維持される作業メモリ表現を更新するとき (およびこの情報が動作を制御するために出力されるとき) を動的な乗算 gating 制御するシステムの計算能力を示します。視床皮質ループに対する大脳基底核の脱抑制(調節)影響は、これと同じ種類のゲート機能を効果的に生成し、PBWM モデルは、位相性 [[dopamine]] 信号が生物学的に妥当な方法でこのゲート機能をどのように訓練できるかを実証しました。
{id=”figure_alm-thal-loop” style=”高さ:25em”}
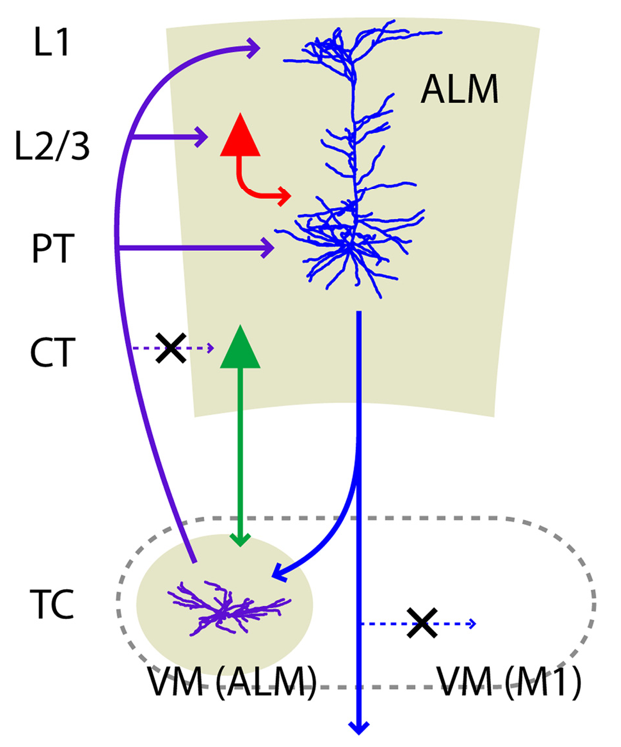
最近の実験データは、PFC の能動的維持に対する視床のこのゲートのような影響を強力に裏付けています。解剖学的観点では、[[@^GuoYamawakiSvobodaEtAl18]] は、[[#figure_alm-thal-loop]] に示すように、複数の高度な神経科学ツールを使用して、ALM 野 (げっ歯類 dlPFC) の第 5b 層 PT (錐体路) ニューロンと VM (腹内側) 視床核の間の視床皮質ループの正確な性質を特定しました。 VM は一次運動野 (M1) にも投射しますが、PFC PT ニューロンから入力を受け取る特定のニューロンも、M1 ではなく ALM に送り返し、興奮性閉ループを形成します。これは、活発な神経発火を維持するために重要であると考えられます。
{id=”figure_economo-18” style=”高さ:30em”}

深層 5 の PT ニューロンの組織のさらなる改良は、[[@^EconomoViswanathanTasicEtAl18]] によって提供され、[[#figure_economo-18]] に示されているように、これらのニューロンの 2 つの異なるタイプの証拠を発見しました。 PT 上部ニューロン (層 5b 上部) は [[#figure_alm-thal-loop]] に示される視床皮質ループに関与するニューロンであり、PT 下部ニューロン (5b 下部) は延髄などの脳幹 [[motor]] 領域に下方に投影されます。
TODO: これを理解してください: L5a IT -> thal、str (?)、L5bU PT -> thal、str (PTp?)、L5bL PT -> subcort
これら 2 つのニューロン タイプの存在により、準備運動計画と実際の運動実行を区別するメカニズムが提供され、計画段階 ([[@ChurchlandShenoy24]]) での時期尚早な実行が防止されます。しかし、[[@^EconomoViswanathanTasicEtAl18]] は、これら 2 つの神経集団の間には単純なきれいな解離が存在せず、両方のタイプのニューロンが遅延期と応答期に活動を示すことを発見しました。それにもかかわらず、PT 下部ニューロンが反応の直前および反応中に発火する可能性が増加しましたが、PT 上部ニューロンは準備遅延期間中に持続的な発火を示す可能性が高くなりました。したがって、[[distributed representations]] で説明し、[[#figure_hunt-rsa]] で示したように、学習によって形成された集団レベルのパターンが常に運転行動に最も関連します。
これらの視床皮質ループと初期の計算モデルの特性と一致して、視床がタスク関連の状況で PFC 活動のダイナミクスを形成する上で実際に重要な役割を果たしていることが複数の研究で示されています ([[@StokesBest90]]; [[@GilmartinBalderstonHelmstetter14]]; [[@GuoLiHuberEtAl14]]; [[@BolkanStujenskeParnaudeauEtAl17]]; [[@SchmittWimmerNakajimaEtAl17]]; [[@DacreColliganClarkeEtAl21]]; [[@WilhelmSychFominsEtAl23]];
{id=”figure_bolkan-17” style=”高さ:15em”}
{id=”figure_guo-14” style=”高さ:15em”}

げっ歯類の視床の MD 核と PL PFC 間のループに焦点を当てた研究が多数あります (総説については [[@ParnaudeauBolkanKellendonk18]] を参照)。たとえば、[[#figure_bolkan-17]] は、比較的長い遅延期間にわたって維持される PL ニューロンの活動を示します。これは、維持のために MD 視床を通るループに依存し ([[@BolkanStujenskeParnaudeauEtAl17]])、腹側海馬からの入力にも依存していました ([[@SpellmanRigottiAhmariEtAl15]])。 [[#figure_guo-14]] に示すように、より短い時間ウィンドウでの同様の結果が [[@^SchmittWimmerNakajimaEtAl17]] および [[@^GuoLiHuberEtAl14]] によっても見つかりました。
これらおよびその他の研究では、PFC で保持されている情報と MD から生じる位相更新信号に基づいて、PFC または MD ニューロンの異なる集団が相互作用を駆動している、課題のさまざまな段階で活動する個別の回路を分析することができました。したがって、初期の計算モデルと一致して、複雑な逐次認知処理には動的な視床皮質相互作用が関与していることは明らかです。
{id=”figure_inagaki-22” style=”高さ:10em”}
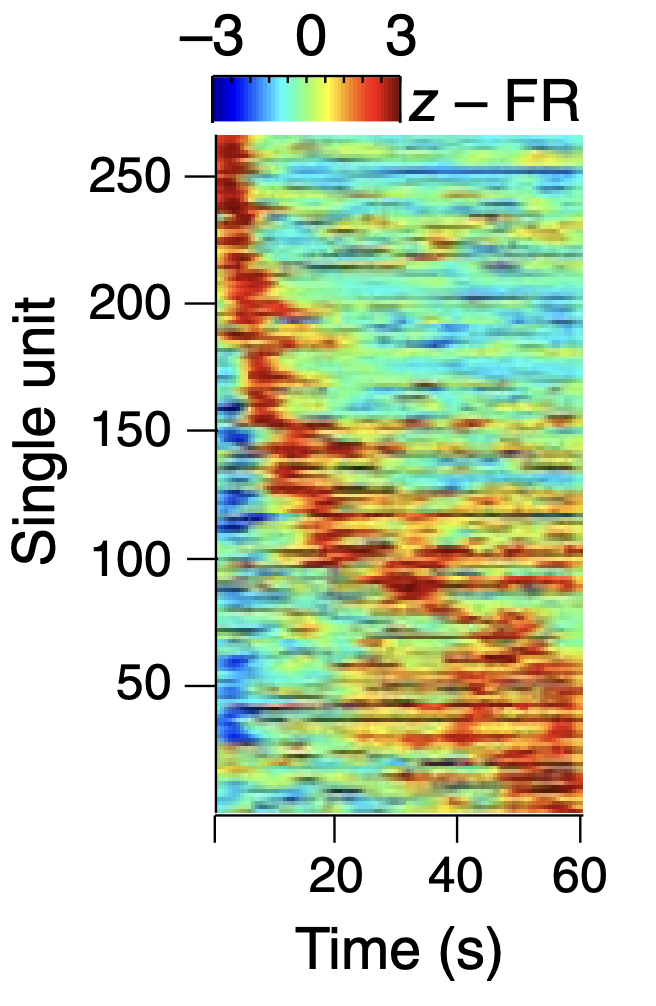
PL_MD 回路に加えて、他の PFC 領域も同様の視床皮質ダイナミクスを持っています。たとえば、[[#figure_inagaki-22]] は、ALM 準備活動によって駆動されるモーター応答をトリガーするための視床入力間のゲートのような関係を文書化した [[@^InagakiChenRidderEtAl22]] の結果を示しています。彼らはこれらの結果を、視床に突き出ているPPN(足橋核)への直接の聴覚入力という観点から解釈している。しかし、PPNは大脳基底核への広範な投射も持っており、これがこの課題の獲得に重要な役割を果たす可能性が高く、SNrニューロンがGoの合図に対して短潜伏の脱抑制反応を示すことが示された。
このタスクが PFC に依存する主な理由は、命令キューと応答するための Go キューの間に遅延があるためです。大脳基底核は、その時間間隔にわたって情報を確実に維持することができません。また、ALM と相互作用する多くの視床領域から記録され、PPN 投影は、目標主導型 PFC 領域 (たとえば、以下および [[thalamus]] で詳しく説明する MD および PF/CM) と強く関連する領域に対して最も強かった。
視床皮質ループの調節における大脳基底核の役割に関するより直接的な証拠は、[[basal ganglia]] でレビューされた広範な文献を含む他の複数の研究から得られます。たとえば、[[@^WilhelmSychFominsEtAl23]] は、作業記憶課題における PL ニューロンから背内側線条体 (DMS) への投射の重要性を文書化しており、[[@^AkhlaghpourWiskerkeChoiEtAl16]] は、[[#figure_bolkan-17]] に示すように、PL ニューロンと非常によく似た DMS ニューロンの遅延期間活動を示しています。
特徴的な視床ゲート回路
{id=”figure_haber-ct” style=”高さ:30em”}
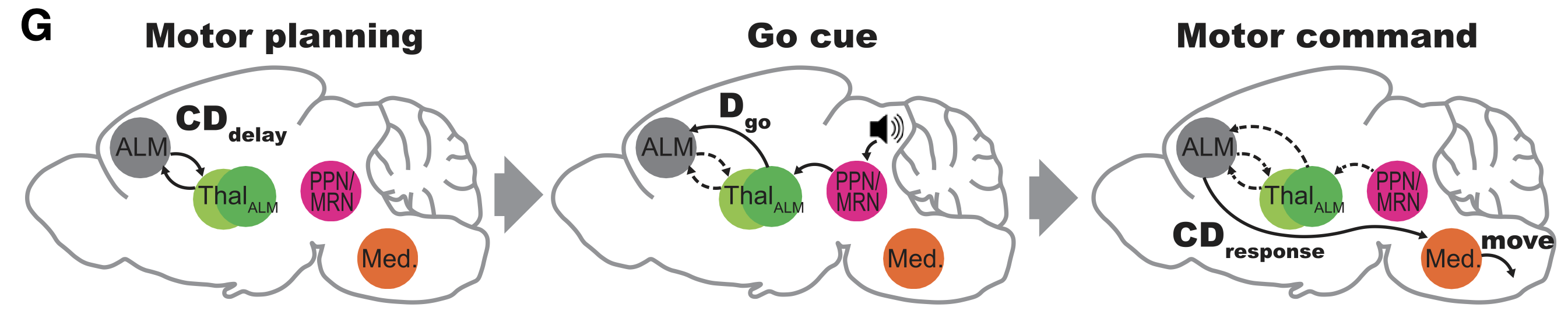
ルビコンの目標主導型フレームワークでは、質的に異なる 2 つのタイプの視床皮質ゲート システム ([[#figure_haber-ct]] から [[@McFarlandHaber02]]) が存在すると仮定しています。
-
ゴールゲーティング: [[#figure_goal-bio]] に示すように、dlPFC 様領域 (げっ歯類の ALM など) も含む、腹側および内側 PFC 領域の分散ゴール領域に対するゲーティング制御を駆動する共通回路が存在する必要があります。視床の MD (中背核) 核の独特な接続性は、まさにこの形式の接続性を提供しており、標準的な BG 出力経路に加えて腹側淡蒼球出力経路を介して、大脳基底核の腹側領域および内側領域からの脱抑制制御下にあります。
-
作業記憶と運動ゲーティング: 関与した目標の文脈内で、運動野は視床投影によって調節され、予期的な運動計画やその他の形態の能動的な作業記憶をサポートするために積極的な維持を維持することができます。腹側領域には、この機能をサポートする他の視床核 (VM、VA、VL など) がいくつかあります。興味深いことに、げっ歯類のALMを含むdlPFC領域は、両方の目標ゲートネットワークに属し、目標を達成するための全体的な運動計画に関与すると同時に、運動計画が時間の経過とともに展開するにつれてより動的に更新されるという点で「デュアルポート」されています。
{id=”figure_motor-thal” style=”高さ:30em”}

これらの異なる視床皮質ループのより詳細な説明は [[thalamus]] ページで提供されますが、主要なポイントは、げっ歯類の MD 野のゴールゲート制御接続を示す [[#figure_md-pfc]] から [[@^KuramotoPanFurutaEtAl17]] と、運動階層に従って編成された運動野視床皮質ループを示す [[#figure_motor-thal]] にまとめられています。 ([[@Bosch-BoujuHylandParr-Brownlie13]])。
MD の場合、内側部分 (MDm) は、目標に関連する PFC 領域のすべてに対してゲート制御の単一の場所を提供し、中央 (MDc) は OFC (結果) 領域へのより具体的な接続を提供し、外側 (MDl) は PL および ACC 領域と接続します。これは、アクティブな目標に取り組むためのグローバルな目標全体のゲート信号と、物事が進むにつれて目標状態の結果および努力/有用性の側面をより具体的に更新する両方が存在するという考えと一致しています。
霊長類特有の PFC 領域
TODO:
- PFC は霊長類、特にヒトで大幅に拡大
*げっ歯類では明らかではない、解剖学的に明確な新しい領域が存在し、主にブロードマン野8、9、46に存在するが、44、45、47にも存在する(口および舌の補助運動野へのIFG)。これらは言語にとって重要です。
- Tom Hazy による最近の Phillips らによる 9/46 dlPFC エリアとの VAmc 接続に関する研究。
ヒトにおけるいくつかの神経画像研究では、大脳基底核と PFC の間の潜在的なゲーティング関係が調査されています。たとえば、[[@^vanSchouwenburgdenOudenCools10]] は、[[fMRI]] の下で [[dynamic causal modeling]] を使用することにより、腹側線条体の活動が dlPFC 野 (特に、人間のタスク表現の維持に関与することが多い IFG) に反映されるタスク切り替えを引き起こし、その後、刺激固有の後皮質領域 (顔やシーン) の活動を駆動することを示しました。おそらく VAmc を駆動するものである DMS の関与を正確に排除することは困難ですが、VS に特有の活動は、MD 視床に投射するこれらの領域が、より集中的な純粋な認知レベルの更新ではなく、タスク切り替えの場合におけるより大規模な目標状態の更新を駆動している可能性があることを示唆しています。その後の研究では、腹側線条体が注意の切り替えの場所であることも発見され([[@vanSchouwenburgdenOudenCools15]])、[[@vandenBoschLambregtsMaattaEtAl22]]は腹側被殻のドーパミンレベルの違いが個人差や反転学習における薬理学的操作に関連していることを示しています。
PFC の計算実装
- PT、PTp は [[@EconomoViswanathanTasicEtAl18]] に基づいて名前変更されますか? ITとの関係は?