compcogneuro/web: space
このページは外部資料の日本語訳です。原文の見出し順と本文順を保ち、コード・URL・出典表記はできるだけ原形のまま残しています。
出典とライセンス
原典: https://github.com/compcogneuro/web/blob/main/content/space.md
ライセンス: Text: CC BY 4.0; code: BSD 3-Clause。このページは日本語翻訳であり、変更点は翻訳とサイト内整形です。
+++ Categories = [“Neuroscience”, “Cognition”] bibfile = “ccnlab.json” +++ 空間の脳の表現は、明確に定義された 1 つの解剖学的システムだけに属さない、相互接続された脳領域のシステムによって確立されます。 [[distributed representations]] の原則に従って、これらのさまざまな領域にわたって空間情報を表現する方法は複数あります。
種が異なれば空間のさまざまな側面が強調され、齧歯動物は特に十分に文書化された一連の航行空間表現を持ち、より大きな環境内を移動する能力をサポートします。霊長類は、柔軟な目と頭のサッカードの動きに基づく、よく発達した眼空間システムを追加しており、これは、さまざまな種類の機会と脅威に満ちたジャングルなどの複雑な環境での視覚探索に使用されます。
モーション (位置の一次導関数) は、これらすべてのスケールにわたる空間処理の大きな部分を占めており、[[vestibular]] 信号は頭と体の動きの感覚表現を提供します。動きはまた、速度と加速度を計算するために [[time]] を暗示します。空間経路のいくつかの重要な要素は、さまざまな方法で時間を表すことに関係しています。
空間表現の構築に関与する領域の多くは [[hippocampus]] システム (およびその一部) と緊密に相互接続されており、これは [[episodic memory]] にとって重要であるため、空間とメモリの関係も重要です。時間と記憶は、空間システムの主要な要素によって生成されブロードキャストされる海馬機能に対する [[theta rhythm]] の強い影響により、速いタイムスケールであっても明らかに強い相互関係があります。
現象学的レベルでは、毎年数千マイルを移動する鳥や魚から、より小規模ではあるが、生息地周辺の動物の知的ルート探索に劣らず印象的なものまで、動物の行動の最も驚くべき例のいくつかは、印象的なナビゲーション能力に基づいています (多くの例については、[[@GrievesJeffery17]] を参照)。
空間参照フレーム
-
他動中心は外部環境に相対的なものです。
-
自己中心的は、あなた自身の身体に関してです。これには、目を中心にした、頭中心にした、体幹を中心にしたなど、さまざまなバリエーションがあります。
パスの統合
経路統合 (および推測航法) は空間処理の 2 つの基本要素であり、さまざまな方向ベクトルに沿った位置の増分変化に基づいて現在地を追跡する機能を反映しています。これらは、速度ベクトルの計算に必要な空間と時間の本質的な統合を表しており、基礎となるニューラル システムの性質を理解するのに役立ちます。
図Xのように、北方向に毎秒10cmの速度で20秒間歩き、その後45度右に曲がり、さらに同じ速度で20秒間進むと、どこに着くか分かるはずです。ご覧のとおり、これらの計算には、ある種の異動中心基準座標系 (たとえば、「北」) に対する運動の角度と運動の速度を知ることが含まれます。
これらの量はどちらも、回転形式と線形形式の加速度 (つまり、位置の二次導関数) を検出する前庭系からの感覚信号に基づいて、内部コンパスのような進行方向と速度の推定値を段階的に更新することによって計算できます。
このような積分を実行するには、加速が発生した時間を何らかの形で表現する必要があります。実際、正確な時間を計ることの重要性は、初期の海洋探検家にとって大きな技術的課題でした。この経路統合プロセスを駆動するのと同じシステムで生成されるシータリズムが、この統合プロセスの基本的なタイミング信号を提供すると思われます。
乳頭視床系
{id=”figure_mmt” style=”高さ:50em”}
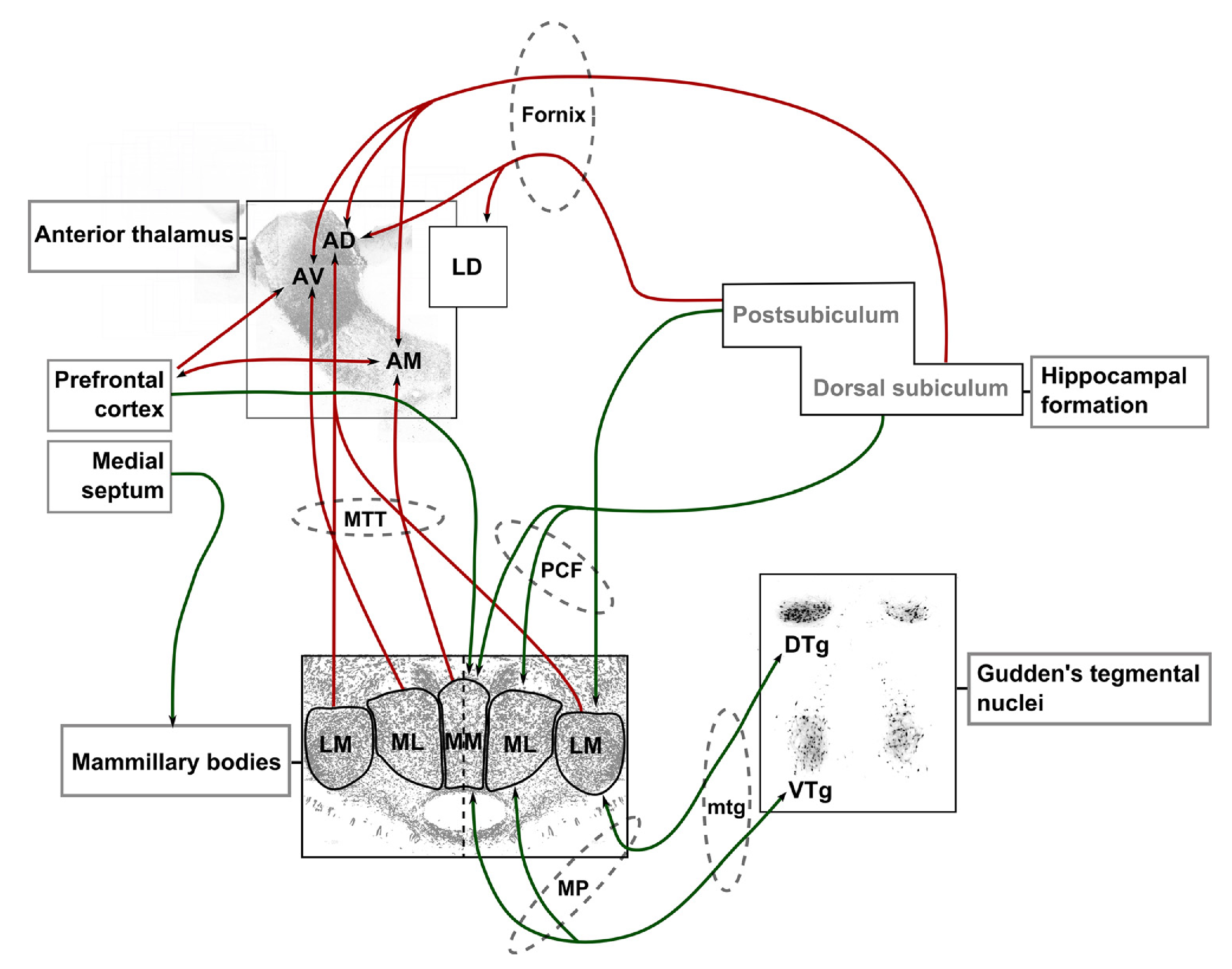
乳頭視床系 ([[#figure_mmt]]) は、経路統合プロセスを固定する [[thalamus]] の前核を中心とした領域のネットワークであり、グッデン被蓋核として知られる中脳領域、具体的には DTN (背側被蓋核) と VTN (腹側被蓋核) で生成される 2 つの基本信号で始まります。核)。 DTN のニューロンのほとんどは、前庭入力に基づいて頭部速度情報をエンコードしますが、VTN のニューロンは、一連の細胞機構によって生成されるシータ タイムスケール (200 ms、10 Hz) での位相的バーストを示します。
DTN と VTN は両方とも、乳頭体標的に抑制性の投射を送ります。たとえば、DTN は乳頭体外側核 (LMN) を抑制します。 LMNは、_頭方向細胞_が最初に始まる場所であり、前背側(AD)視床核を介した回帰システムを介して背側前台(後台としても知られています)に投射し、その後LMNに戻って投射します(図)。
頭方向セル
LMN における頭部方向細胞の出現は広範囲に研究されており、比較的よく理解されています ([[@Taube07]])。重要な計算メカニズムは、ほぼ同時期に発表された一連の 4 つの基礎論文 ([[@SkaggsKnierimKudrimotiEtAl94]]、[[@SharpBlairBrown96]]、[[@Zhang96]]、[[@RedishElgaTouretzky96]]) によって提案されているように、頭部の 360 度のコンパスのような方向を異中心座標で表す リング アトラクター です。
isocoding ニューロン (同じ方向をエンコードするニューロン) 間の興奮性ループは、現在の頭部方向信号を維持しながら、他の方向を横方向に抑制します。これは、複数の脳領域にわたる [[stable activation]] の標準レシピです。 [[Vestibular]] 信号は頭の動きを反映し、この活動の「隆起」を周囲に押し出します。
このようなシステムの主な問題点は、適切に校正された信号を生成するためにシナプス ウェイトを正確に調整する必要があり、ヘッドが動いていないときに信号が長時間にわたって安定していることです。リングアトラクターは、脳内に遍在するノイズ ([[@BrodyRomoKepecs03]]; [[@BarakTsodyks14]]; [[@LimGoldman14]]) による摂動に敏感であることで知られています。
[[vestibular]] システムにおけるこのような微調整の問題に対する明確な解決策は、主に他の感覚信号に対する自己の動きの影響を正確に予測してキャンセルすることを学習する [[cerebellum]] によって提供されます。この小脳の学習の重要な結果の 1 つは、頭の方向の更新を駆動する入力には、頭と体を動かすあらゆる種類の運動動作の影響が自動的に含まれており、小脳はその影響を前庭の影響についての対応する感覚予測に統合することです。
小脳内のこの同じエラー駆動型 [[predictive learning]] メカニズムは、リング アトラクター シナプスを調整して、入力がない場合でも一貫した頭の方向のバンプを正確に維持し、正味の頭と体の動きを反映するように正確に更新することもできます。