compcogneuro/web: synaptic-plasticity
このページは外部資料の日本語訳です。原文の見出し順と本文順を保ち、コード・URL・出典表記はできるだけ原形のまま残しています。
出典とライセンス
原典: https://github.com/compcogneuro/web/blob/main/content/synaptic-plasticity.md
ライセンス: Text: CC BY 4.0; code: BSD 3-Clause。このページは日本語翻訳であり、変更点は翻訳とサイト内整形です。
+++ Categories = [“Learning”, “Neuroscience”] bibfile = “ccnlab.json” +++ シナプス可塑性は、シナプス接続の強さ (つまり、重み) が変化するという確立された生物学的現象であり、これは脳における学習の主な基礎であることが知られており、[[abstract neural network]] モデルの基礎仮説と一致しています: 知識はシナプスの重み_にエンコードされている。
-
LTP = 長期増強 は、[[@^BlissLomo73]] によって最初に確立された、神経活動の結果としてシナプス強度が持続的に増加する現象の名前です。
-
LTD = 長期うつ病は、シナプス強度の_減少_を指し、確立するまでにしばらく時間がかかりました ([[@BearMalenka94]])。
これらの現象の生化学的基礎を明らかにすることは、神経科学における最も成功した共同研究プロジェクトの 1 つであり、その結果、ここで簡単にレビューするように、関係する経路やプロセスの多くがさまざまな時間スケールで展開する詳細な理解が得られました。
| 理想的な世界では、この取り組みの結果は、[[neocortex]] での学習の_機能的_性質の完全なボトムアップの基礎となるでしょう。つまり、基礎となるハードウェアの完全な理解に基づいて、アルゴリズム的および計算的な [[levels of analysis | level of analysis]] で_学習アルゴリズム_ を導出できるはずです。 |
根底にある神経生物学的メカニズムからこの学習アルゴリズムを導き出そうとする既存の試みは、Donald Hebb ([[@Hebb49]]) によって先見的に説明された [[Hebbian learning]] の原理を中心にしています。ここでの核となるアイデアは、「一緒に発火するニューロン、一緒に配線する」という言葉に込められています。言い換えれば、ヘビアン学習は、神経活動のパターンが時間内に同時発生する限り、何らかの形で互いに関連するニューロン間の接続を構築する、連想型の学習形式です。結果として得られる連想ネットワークは、私たちが世界を主観的に概念的に理解するための適切な基盤を提供するように思えます。たとえば、煙を見ると、私たちは火の概念を連想し、したがって有用な推論を行うことができます。
[[#spike timing dependent plasticity]] (STDP) ([[@BiPoo98]]) の現象は、ヘビアンの考え方にさらなるニュアンスを加え、LTP と LTD の両方を 1 つの一貫したフレームワークにエレガントに統合します。ニューロンは明らかに、発火の時間的順序にも敏感であり、シナプス前ニューロンがシナプス後ニューロンより前に発火する場合は連合強化 (LTP) が、逆の場合は連合弱化 (LTD) が起こります。直観的には、これらのタイミング関係が因果的な順序を表す場合、これによりニューロンは「因果的」種類の関連性を学習することができます。非常に一般的に言えば、シナプス前ニューロンがシナプス後ニューロンよりも先に発火する場合、それがシナプス後ニューロンの発火に寄与した可能性が高くなります。
しかし、この非常に局所的な神経発火因果関係が、より大きな環境因果関係と関係があるという考えには、さらに多くの仮定が必要であり、それを維持するのは困難です。さらに、広範な追加研究により、STDP 現象は、覚醒して行動している状態 ([[in activo]]) ([[@DebanneInglebert23]]; [[@ShouvalWangWittenberg10]]; [[@FroemkeTsayRaadEtAl06]]; 重要なイオンのより正確な生理学的レベルを含む: [[@InglebertAljadeffBrunelEtAl20]]) におけるより自然な神経活動パターンに一般化されない、非常に特殊な人工条件で発生することが示されています。
[[Hebbian learning]] で説明したように、連想学習原理には (STDP バージョンであっても)、[[neocortex]] の認知機能を達成できるニューロンの深層ネットワークの効果的な全体的な組織化がもたらされるという考えを裏付ける数学的または計算的基盤がありません。これらすべての考察により、ヘビアン学習はシナプス可塑性の完全な計算レベルの記述を提供できないという結論につながります。
代わりに、[[error-driven learning]] を [[predictive learning]] と矛盾しない方法で実装するシナプス可塑性の一貫した計算レベルの記述を提供するには、異なる時間ダイナミクス (対 STDP) が重要であると主張します。ヘビアン学習とは異なり、エラー駆動学習は、現在の [[artificial intelligence]] (AI) の進歩を支えるすべての最新の [[abstract neural network]] モデルに確立された基盤を提供するため、計算レベルで機能することがわかっています。さらに、予測学習フレームワークは、ヘビアン連想学習フレームワーク、特に STDP 因果学習とほぼ同じ直観的理解を捉えています。新皮質における学習の主な推進力は、次に何が起こるかを予測することです。これは、煙と火の連想学習の例についての別の考え方です。
既知のシナプス可塑性メカニズムを使用してエラー駆動学習を実装するために必要な重要な時間ダイナミクスは [[temporal derivative]] です。これは、異なる [[exponential integration]] 時定数を持つ神経活動を統合する 2 つのプロセス間の競合相互作用を介して計算できます。これは、[[Axon]] で使用される [[kinase algorithm]] の基礎であり、シナプス可塑性 (LTP 対 LTD) の方向を決定する 2 つのキナーゼ間の競合的相互作用にちなんで名付けられました ([[@GoodellZaegelCoultrapEtAl17]])。このアルゴリズムは、以下で説明するすべてのメカニズムに基づいて構築されています。
シナプス可塑性メカニズム
{id=”figure_synapse” style=”高さ:30em”}
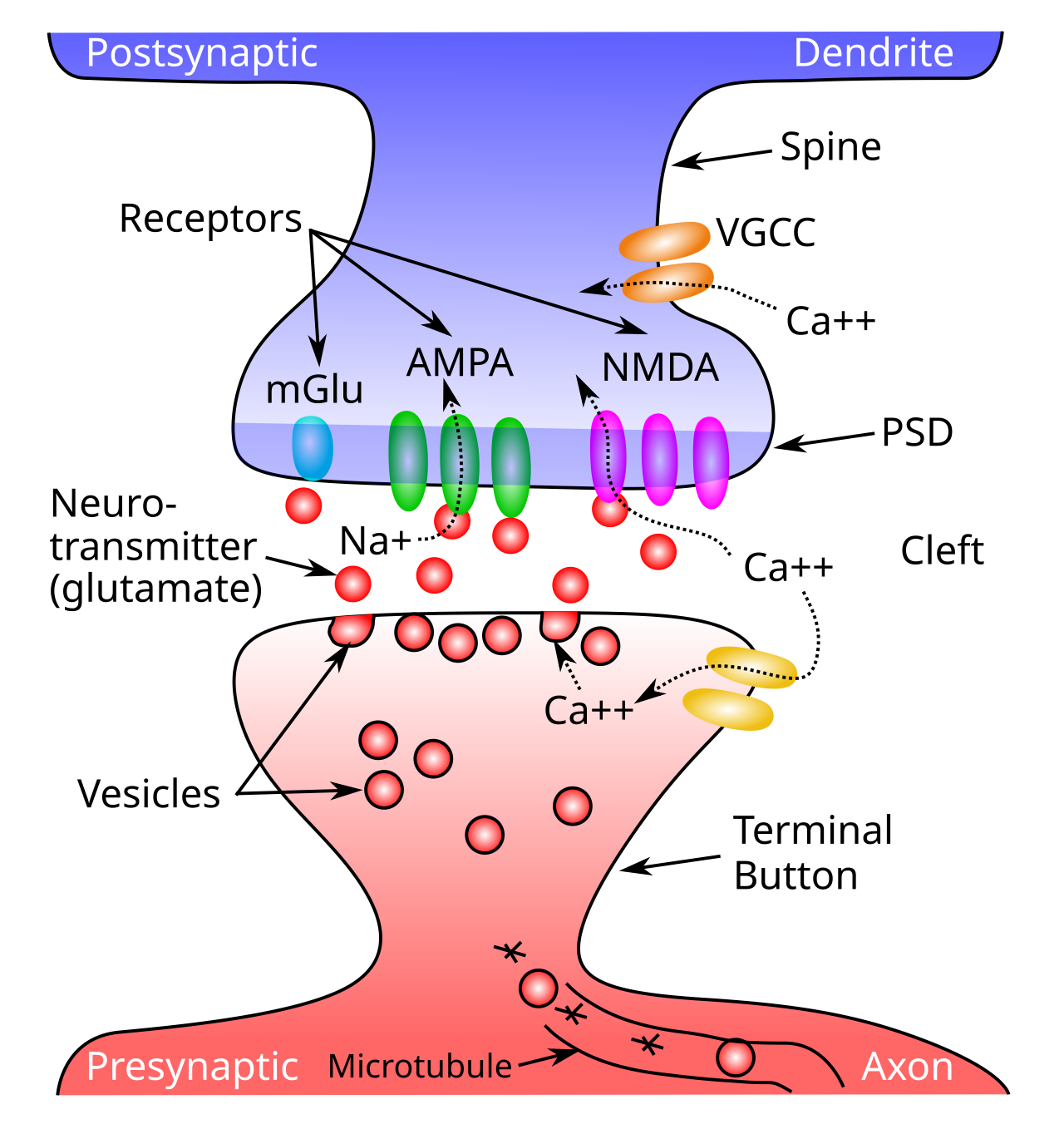
シナプス ([[#figure_synapse]]) には多くの可動部分 ([[neuron]] で紹介) があり、そのうちの 1 つが全体の効率を変化させる重要な要因となる可能性があります。あなたはいくつ思いつきますか?重要な因子の探索は、シナプス可塑性に関する研究の初期段階を支配しており、放出されるシナプス前神経伝達物質の量から、シナプス後AMPA受容体の数と有効性、さらにはシナプス前成分と後成分の整列などのより微妙なものや、複数のコピーへのシナプス接続のクローニングなどのより劇的な変化に至るまで、さまざまな因子の関与の証拠が長年にわたって発見されてきました。
私たちの現在の理解では、[[neocortex]] の興奮性ニューロンにおけるシナプス可塑性の支配的な要因は、活性シナプス後 AMPA 受容体の数と有効性であるということです。AMPA 受容体は、シナプス後密度 (PSD) に埋め込まれており、グルタミン酸が結合できるようにシナプス間隙に露出している受容体です。アクティブな AMPA チャネルの数は、内部の「予約」ストアと PSD の場所の間でこれらのチャネルをアクティブに移動する trafficking プロセスによって変更されます。 AMPA 受容体を PSD 内に押し出すには、エキソサイトーシス のプロセスが含まれ、一方、活性な AMPA 受容体の除去には、エンドサイトーシス が含まれます。これらは、すべての細胞で驚くほど速いタイムスケールで行われる日常的なプロセスです。細胞膜を、容易に浸透できる非常に柔軟な脂肪物質としてイメージすることが重要です。
LTP と LTD が生化学レベルでどのように機能するかを理解するには、この AMPA 輸送プロセスを最終的に制御する一連のプロセスを理解する必要があります。これらは異なる段階にグループ化できます。
-
誘導 はすべてを開始する最初の段階です。シナプス前およびシナプス後ニューロンの活動の影響を受ける、ミリ秒単位のタイムスケールで行われる最初のプロセスです。これは、結果が LTP か LTD (または変化なし) になるかを決定する必要があり、生物学的学習アルゴリズムの性質を決定するための重要な段階です。したがって、ここではこれが私たちの議論の焦点です。
-
発現は、誘導プロセスによって引き起こされるシナプス効率の初期変化を伴う次の段階です。これには、PSD 内の既存の活性 AMPA 受容体のコンダクタンスの非常に急速な変化と、活性 AMPA 受容体の数の変化をもたらす AMPA 受容体輸送の変化の両方が含まれます。さらに、樹状突起スパインの「構造」の急速な変化により、この輸送プロセスが促進されます ([[@YangLiu22]]; [[@BoschCastroSaneyoshiEtAl14]]; [[@MatsuzakiHonkuraEllis-DaviesEtAl04]])。この構造は、筋肉に含まれる同じ繊維であるアクチン ポリマー繊維によって部分的に決定されます。
-
早期メンテナンス (E-LTP という用語に含まれる) は、これらの発現変化を数秒から数分にわたって維持するメカニズムであり、この段階で、その変化がベースラインまで減衰して最終的に 短期 可塑性 (STP) になるか、次の段階まで持続する 長期 可塑性になるかを決定します。これらのメカニズムは、起こった発現の変化を「安定化」させる必要があり、細胞の内部は複雑な化学反応が連続的に起こる非常に混沌とした活動的な場所であるため、この早期の維持は初期の発現変化の自動的な結果ではないことを理解することが重要です。この早期維持は、GluN2B ([[@BayerGiese25]]) として知られる NMDA 受容体の特定の位置にある NMDA 受容体への_CaMKII_ 分子の結合と、その結果として生じる脊椎の形状の構造変化との相互作用 ([[@YasudaHayashiHell22]]) に依存しているという証拠があります。
-
後期メンテナンス (L-LTP) は、タンパク質合成 (新しいタンパク質分子の作成) を含む最初の段階であり、これには RNA と リボソーム の協調的な相互作用が必要であり、通常、ニューロンでの誘導後約 60 分かかります。新しいタンパク質は PSD ([[@TanakaHoriikeMatsuzakiEtAl08]]) に局在しており、脊椎のアクチン線維構造の持続的な変化にも関与しています。
-
睡眠プロセス: 睡眠中にさまざまな変化が起こり、日中に起こったシナプスの変化をさらに安定させ、更新します。睡眠中に体全体で大きな再構築と構造変化が起こり、脳のシナプスも例外ではありません。一般的な考えの 1 つは、シナプスの全体的な強度が睡眠中にさまざまな方法で再調整されるというものであり ([[@AbbottNelson00]]; [[@TononiCirelli03]]; [[@Turrigiano08]]; [[@TorradoPachecoBottorffGaoEtAl21]])、シナプスの強度がさらに強化されるという証拠もあります ([[@LiMaYangEtAl17]])。これらの変化はすべて、十分に文書化されている睡眠後に起こる記憶の変化に寄与する可能性があります ([[@KlinzingNiethardBorn19]])。 [[@^TorradoPachecoBottorffGaoEtAl21]] で説明されているバージョンのメカニズムを組み込むと、モデルの全体的な安定性と有効性が大幅に向上することがわかりました ([[kinase algorithm#Slow weights]] を参照)。
このページの残りの部分では、誘導プロセスに関与するメカニズムと、それらが残りの段階とどのように相互作用するかに焦点を当て、これらの他の段階の詳細についてはこれ以上議論しません(最近のレビューについては上記の参考文献を参照してください)。注目すべきことに、誘導プロセスの中心人物である_CaMKII_は、その後の段階([[@BayerGiese25]]; [[@YasudaHayashiHell22]])への重要な橋渡しをしていますが、ジョン・リスマンによって提案された元の仮説とは大きく異なる方法ではありますが、その方法には、CaMKII自体が_自己リン酸化_され、それによって時間の経過とともに記憶信号を維持する能力が含まれていました。 ([[@Lisman94]])。むしろ、現在では、[[@^VanHarreveldFifkova75]] によって最初に提案されたアクチン線維とシナプススパインの構造変化が重要な関与をしているため、記憶維持は筋力維持に似ているようです。
NMDA とカルシウムの流入
[[neuron channels#NMDA]] 受容体は、シナプス可塑性の誘導において中心的な役割を果たします。その開口部によってカルシウム イオン ($Ca^{++}$) がシナプス後脊椎に進入できるため、これが AMPA 受容体のコンダクタンスと数の変化をもたらす一連の生化学的カスケードの最初のトリガーとなります。体全体でカルシウムイオンは同様の役割を果たしており、同様の生化学的カスケードが多くの異なる細胞型で見られます。電位依存性カルシウム チャネル (VGCC) もカルシウムを伝導し、代謝型グルタミン酸受容体 (mGluR) と同様に、さらに重要な役割を果たします。
{id=”figure_ltp-nmda” style=”高さ:30em”}
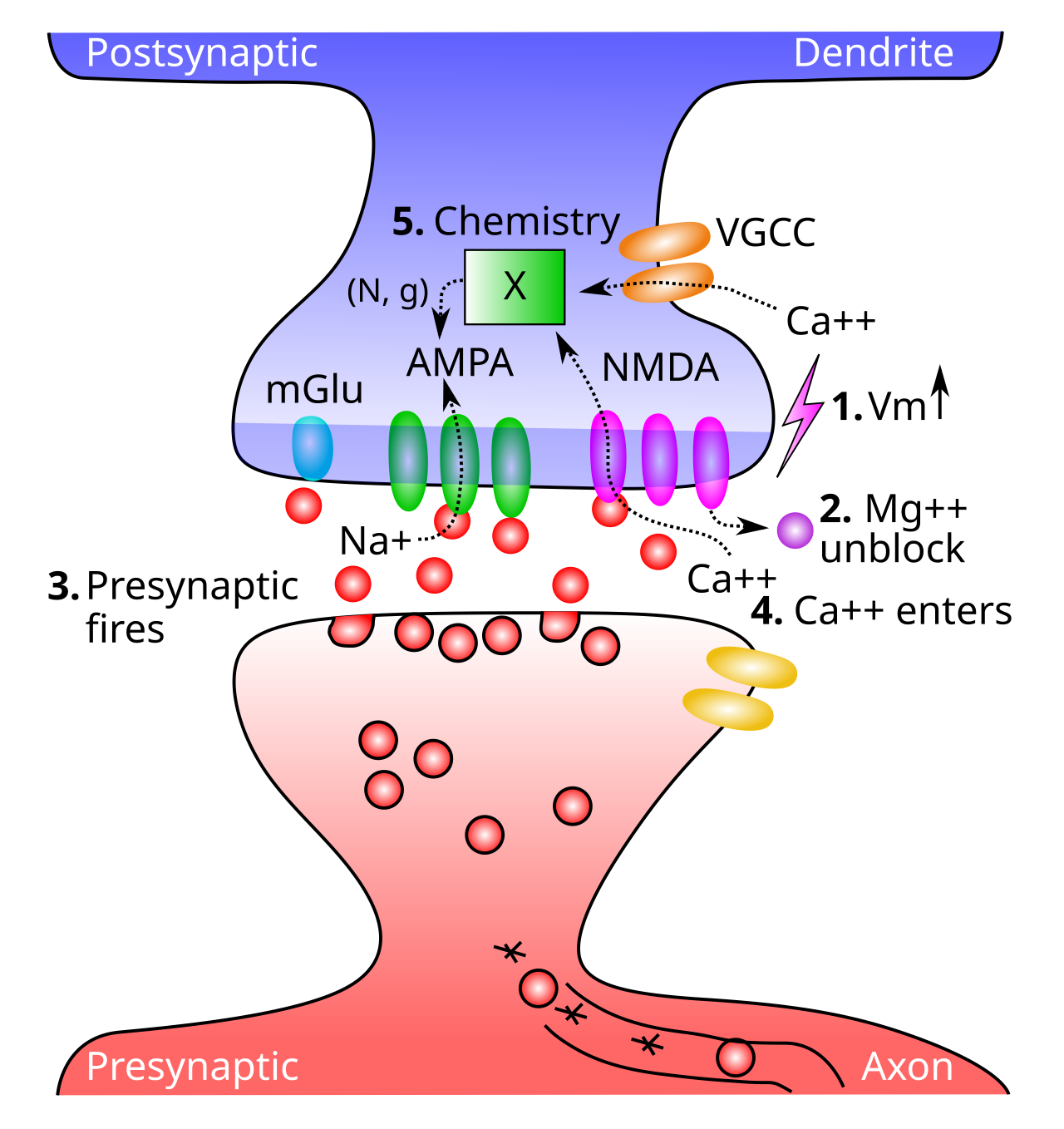
[[#figure_ltp-nmda]] は、NMDA チャネル機能に関係する 5 つの重要なステップを示しています。
-
すべての興奮性シナプス入力が細胞に流入するため、シナプス後膜電位 ($V_m$) が上昇する必要があります。この $V_m$ レベルの最も重要な要因は、実際には 逆伝播する活動電位 です。ニューロンが活動電位を発火させると、活動電位は軸索から前方に進むだけでなく、(樹状突起に沿ったアクティブな電圧感受性の $Na^+$ チャネルを介して) 樹状突起に沿って後方にも下降します。したがって、ニューロン全体がいつ発火したかを知ることができます。これが計算上非常に役立つことがわかります。
-
$V_m$ が上昇すると、マグネシウム イオン ($Mg^{++}$) が NMDA チャネルの開口部から反発され (正電荷が互いに反発し)、チャネルのブロックが解除されます。
-
シナプス前ニューロンは活動電位を発火させ、グルタミン酸神経伝達物質をシナプス間隙に放出します。
-
グルタミン酸は NMDA 受容体に結合し、NMDA 受容体を開いて $Ca^{++}$ イオンがシナプス後細胞に流入できるようにします。これは、NMDA もブロック解除されている場合にのみ発生します。後で説明するように、シナプス前およびシナプス後活動の両方に対する NMDA の依存性は、学習の性質を知るための初期の重要な手がかりの 1 つでした。
-
シナプス後脊椎における $Ca^{++}$ の濃度は複雑な化学反応を引き起こし、最終的に AMPA 受容体の数と有効性を変化させます。これらの AMPA 受容体はニューロンに一次興奮性入力駆動を提供するため、AMPA 受容体を変更すると、シナプス後ニューロンに対するシナプス前活動電位の正味の興奮効果が変化します。これが、シナプスの有効性、つまり_weight_を変更することを意味します。
NMDA はシナプス前部とシナプス後部の両方の活動を必要とするという事実により、NMDA は本質的にヘビアン学習原理の直接的なメカニズムとなり、シナプス前部とシナプス後部の両方のニューロンが同時に活動している場合にのみシナプス接続が増加します ([[@LevySteward79]]、[[@CollingridgeKehlMcLennan83]]、[[@CollingridgeBliss87]])。これが、ほとんどの神経科学者がヘブ学習がすべての話であると自然に想定した理由です ([[@Cooper05]]) — この複雑な生物学的メカニズムが機能的に動機付けられた学習アルゴリズムをこれほど直接実装しているのは単なる偶然でしょうか?
$Ca^{++}$ は、膜電位が上昇した場合にのみ開くカルシウム チャネルである 電位依存性カルシウム チャネル [[neuron channels#VGCC]] を介してシナプス後細胞に侵入することもできます。ただし、NMDA とは異なり、それらはシナプス前神経活動に敏感ではなく、シナプス後活動にのみ依存します。後で説明するように、これは計算上重要な意味を持ちます。 VGCC は NMDA よりも $Ca^{++}$ レベルへの寄与が少ないため、依然として NMDA が支配的なプレーヤーです。
代謝型グルタミン酸受容体 (mGluR) もシナプス可塑性において重要な役割を果たします。これらの受容体は、イオンが膜を越えて流れることを許可せず(つまり、イオン向性ではありません)、代わりに神経伝達物質が結合すると直接化学反応を引き起こします。これらの化学反応は、$Ca^{++}$ によって引き起こされる AMPA 受容体の変化を調節する可能性があります。
カルシウム濃度により LTP と LTD が決まります
生物学的学習アルゴリズムの性質を決定するには、シナプス前およびシナプス後の活動の性質に応じて、NMDA および VGCC チャネルからのこのカルシウム信号がどのように LTP と LTD をもたらすかを理解する必要があります。長い間、研究者は主に LTP (一般に誘発が容易である) に焦点を当てていましたが、最終的にはシナプス可塑性の両方の方向が学習にとって同等に重要であることに気づきました ([[@DunwiddieLynch78]]; [[@Lisman89]]; [[@BearMalenka94]])。
重要な発展は、可塑性の方向は細胞に入るカルシウムの「量」に依存し、それはNMDA受容体によって測定される全体的な「シナプス活性」の関数であるという考えでした。 NMDA 受容体の開口は、シナプス前発火速度 (グルタミン酸結合の場合) とシナプス後発火速度 (マグネシウムのブロック解除の場合) の両方に依存するため、Ca++ 流入の総量は、これらの変数とこの活動の全体の継続時間の関数になるはずです。
{id=”eq_srt” title=”シナプス活動 = 送信者 * 受信者 * 時間”} \([Ca^{++}]_i = S R T\)
ここで、$[X]_i$ は、何かの細胞内濃度の標準表記です。
{id=”figure_ltpd-ca” style=”高さ:10em”}
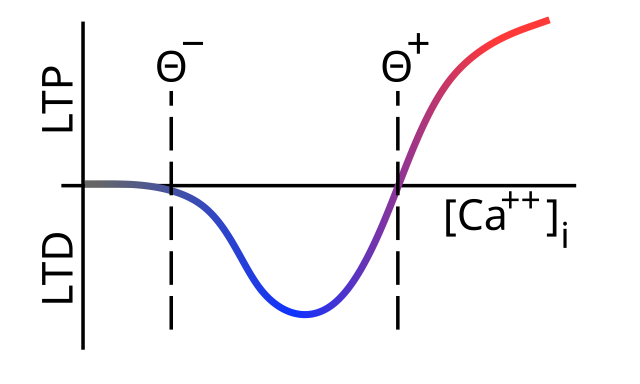
[[#figure_ltpd-ca]] は、変化の方向が少なくとも数百ミリ秒にわたって蓄積されたシナプス後脊椎の $[Ca^{++}]_i$ の全体的なレベルにどのように依存するかについて、今日まで広く受け入れられている標準モデルを示しています。低レベルは LTD を駆動し、高レベルは LTP ([[@Lisman89]]; [[@BearMalenka94]]) を生成します。興味深いことに、[[#figure_ltpd-ca]] の関数の形状は、作成者のイニシャル ([[@BienenstockCooperMunro82]]) にちなんで [[Hebbian learning#BCM]] ルールとして知られる [[Hebbian learning]] のバージョン用に開発されたものと本質的に同一です。
生化学レベルでは、[[@^Lisman89]] の基本モデルは、CaMKII キナーゼが LTP を駆動し、PP1 として知られる競合する_ホスファターゼ_ 経路が LTD を駆動し、$Ca^{++}$ の流入に対するこれら 2 つの経路の感受性の違いにより、図に示す曲線の形状が得られるというものでした。 [[#figure_ltpd-ca]]。具体的には、PP1 は低カルシウム レベルで優勢であるため、LTD を促進しますが、CaMKII は高レベルで優勢であるため、カルシウム レベルが増加するにつれて LTP を促進します。この元の仮説は、CaMKII の機能に関する理解における最近の変化を考慮して大幅に更新する必要があります。これについては、STDP についての以下の説明の後で詳しく説明します。
スパイクのタイミングに依存する可塑性
{id=”figure_stdp” style=”高さ:20em”}
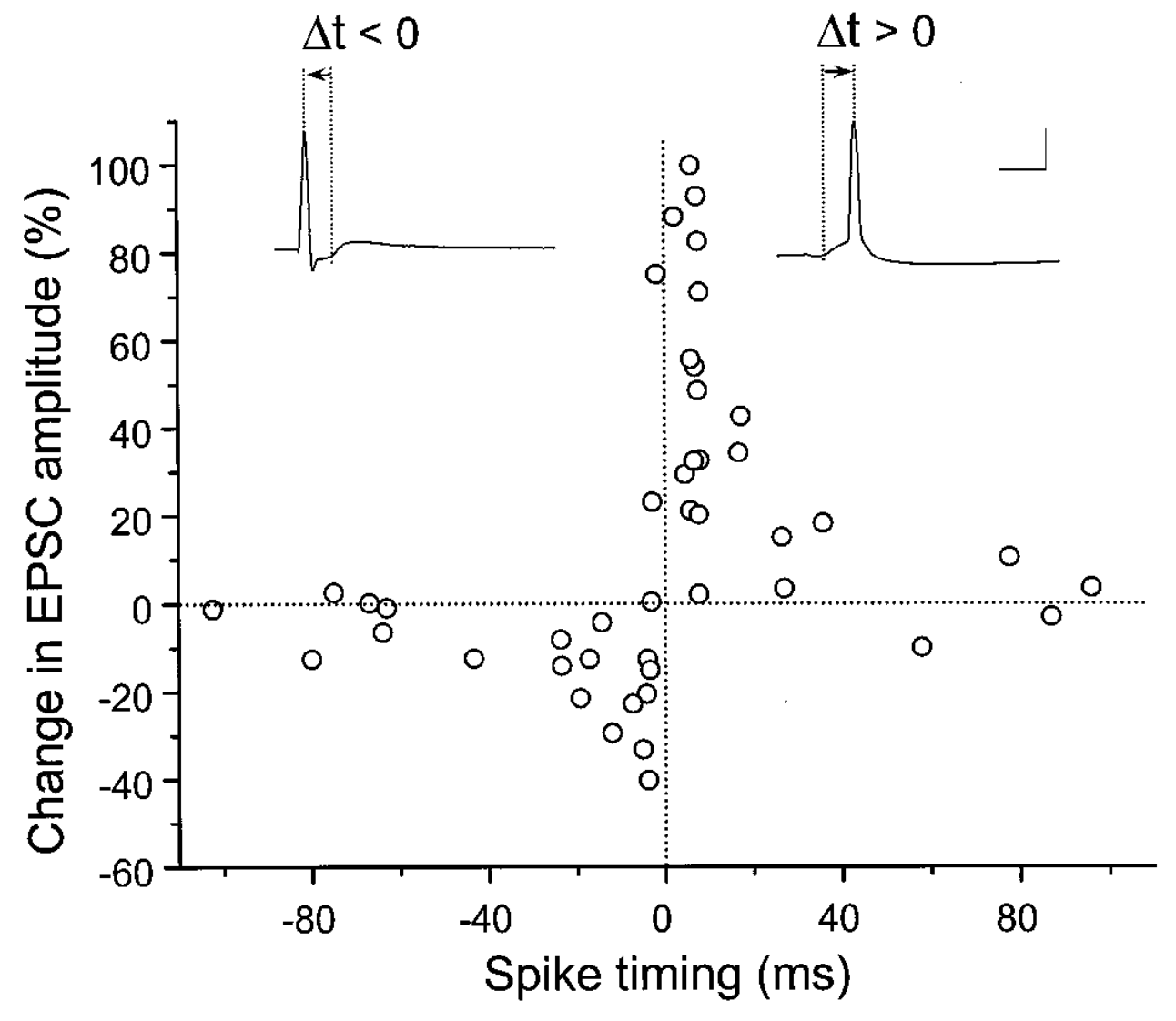
[[@^BiPoo98]] によるスパイクタイミング依存可塑性 (STDP) の発見は、元の [[@^BlissLomo73]] の発見以来、この分野で最も影響力のある唯一の実験であると言っても過言ではありません。グーグル学者 (引用符内!) にその用語が含まれている論文が 30,000 以上あり、元の論文の引用数が 5,000 以上あります。 [[#figure_stdp]] は重要な結果を示しています。シナプス学習は、シナプス前ニューロンがシナプス後ニューロンより前に発火するときに LTP が発生し、逆の順序の場合はその逆であるという、単純で魅力的な 因果関係 原則に従っているようです。この直感的に魅力的なロジックは、STDP のようなメカニズム ([[@KheradpishehGanjtabeshThorpeEtAl18]]、[[@DiehlCook15]] など) を広範囲に採用している多くの計算モデラーにとって特に魅力的であることが証明されています。
因果関係の議論の単純なバージョンは、特定のプレ ニューロンが実際にポスト ニューロンを発火させる役割を果たしている場合、必然的にその前に発火しなければならず、STDP の結果によれば、その重みの強度が増加することを示唆しています。一方、シナプス後細胞の発火に因果関係のない前ニューロンの重みは減少します。
その後、STDP に関する膨大な文献により、in vitro 実験で使用された非自然的なカルシウムおよびマグネシウム濃度への依存性を含む、多くのさまざまな変数 ([[@DebanneInglebert23]]) に応じて、LTP になるか LTD になるか、あるいはどちらにもならないかが非常に大きく異なることが明らかになりました ([[in activo]] を参照)。さらに、元の実験では、STDP の結果を生成するには、1 秒間活動がない状態で区切られた単一の孤立したスパイクのペアを含む、非常に特異的で人工的な刺激条件が必要でした。また、そのようなタイミング パターンを何度も繰り返す必要がありました。
重要なことに、わずかに複雑なスパイク列を使用した場合でも、結果は古典的な STDP 塑性関数の単純な適用と一致せず、代わりに、基礎となるメカニズムの性質がかなり複雑であることが明らかになりました ([[@FroemkeDan02]]; [[@FroemkeTsayRaadEtAl06]]; [[@UrakuboHondaFroemkeEtAl08]])。 [[@^ShouvalWangWittenberg10]] も同様の結論に達しており、BCM ([[@BienenstockCooperMunro82]]) メカニズムのバージョンが全体的なアカウントをより適切に提供すると主張しています。我々は、[[@^UrakuboHondaFroemkeEtAl08]] の生物学的に詳細なモデルを使用してこの結論を再現します。このモデルは、元の人工刺激条件下で古典的な STDP 曲線を生成しますが、以下および [[urakubo08 simulation]] で説明するように、より一般的には BCM のような機能を生成します。要するに、この研究は、STDP が一般的に何を達成するかを適切に説明したものではなく、基礎となるシナプス可塑性メカニズムの特殊な場合の結果であることを示唆しています。
最後に、覚醒しているときの神経活動の自然なパターン、つまり [[neocortex]] の行動を考察すると、そもそも STDP 理論の背後にある因果論理についての根本的な疑問が生じます。具体的には、新皮質のすべてのレベルでかなりの継続的な神経活動が存在しており、これは、たとえば、入力刺激に対するニューロンの応答を強力に文脈化することが示されています ([[@ArieliSterkinGrinvaldEtAl96]])。したがって、STDP に基づくモデルによって想定される厳密なフィードフォワード因果方向とは反対に、脳内の活動には、広範な [[bidirectional connectivity]] のため、単純で信頼できる因果関係の矢印がありません。
キナーゼとホスファターゼの生化学的カスケード
LTP と LTD が脳内でどのように機能するかをより深く理解するには、シナプス後脊椎へのカルシウムの流入によって引き起こされる複雑な生化学的カスケードについてさらに理解する必要があります。まず、生化学の標準原理と、2010 年代を通じて開発された多くの実験結果と理論に基づいた、現在「古典的」モデルと呼ぶことができるものについて説明します。次に、利用可能なデータ ([[@BayerGiese25]]) に全体的によりよく適合する新しい structural モデルについて説明します。
これらの生化学的カスケードには (カルシウム自体に加えて) 2 つの主要なプレーヤーがいます。
-
キナーゼは、他の化学反応を促進するための「触媒」として機能するタンパク質構造です。これらは通常、特定の結合部位で他の物質が結合することによって活性化されます。特定の結合部位は、アミノ酸と、タンパク質鎖の N 末端からのアミノ酸単位の距離を示す番号 (例: Thr286) によって識別されます。キナーゼは通常、他の分子を「リン酸化」するように作用し、それによって分子が「活性化」され、他の分子に結合したり、他の機能を実行したりできるようになります。リン酸化とは、リン酸基 ($PO_4$ に関与する化合物) を追加することを意味します。高校の生物では、ATP が細胞内のエネルギー伝達体であり、ATP の P がリン酸 (アデノシン三リン酸) であることを覚えているかもしれません。リン酸 = 非常に一般的な意味でのエネルギーまたは活性化。
-
ホスファターゼはキナーゼとは反対の役割を果たします。他の分子を脱リン酸化するように作用するため、通常はそれらを不活性化または阻害します。ただし、たとえば分子全体の形状の対応する変化により、1 つの部位の脱リン酸化により実際に他の部位での結合が可能になる場合もあります。したがって、キナーゼは常に興奮性または活性化し、ホスファターゼは常に抑制性または非活性化するという厳密な規則はありません。
{id=”図_ウラクボ-ケム”}
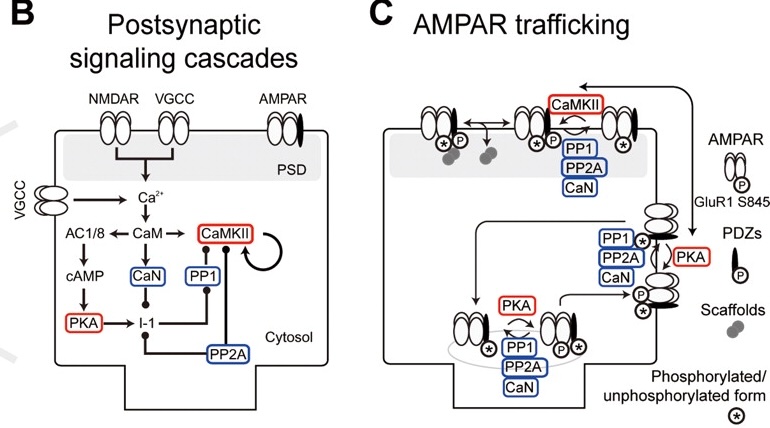
[[#figure_urakubo-chem]] は、NMDA および VGCC 受容体を介した $Ca^{++}$ の流入によって引き起こされる化学プロセスのカスケードの「標準モデル」を示しており、2000 年代の最初の 20 年間にわたる一般的な理解レベルを反映しています。これらすべての化学プロセスについて経験的に決定された速度定数に直接基づいた詳細な生化学モデルが [[@^UrakuboHondaFroemkeEtAl08]] によって開発され、その複製が [[urakubo08 simulation]] で探索可能です。このモデルには AMPA 受容体輸送メカニズムが含まれているため、これらの遅いプロセスが効果を発揮するまでの時間が経過した後、PSD 内の AMPA 受容体の数の観点から、このモデルにおけるシナプス効率の変化を直接測定します。このモデルにはいくつかの重要な新しい要素が欠けていますが、更新されたモデルで使用できる十分に検証されたコンポーネントが多数含まれており、これらの化学プロセスがどのように機能するかを詳細に理解するための有用なツールを提供します。さらに、これから説明するように、広範囲の既存データが考慮されます。
このカスケードは、$Ca^{++}$ の主要な直接結合標的である CaM (calmodulin) から始まり、他の標的への結合を可能にする形状変化を誘導することでこのタンパク質を活性化します。活性化された CaM は、CaN (カルシニューリン、ホスファターゼ) と CaMKII キナーゼ (カルシウム/カルモジュリン依存性プロテイン キナーゼ 2) を活性化します。これについては、すでに何度か言及しましたが、明らかに、シナプス可塑性に関与する相互作用プレーヤーの複雑なネットワークの中で唯一最も重要なタンパク質です。カスケードのさらに下流には PP1 (プロテイン ホスファターゼ 1) があり、[[@^Lisman89]] の標準 LTP / LTD モデルによれば、これは CaMKII の主要な敵対者です。 PKA (プロテインキナーゼ A) という名前の別のキナーゼは、PP1 活性化の阻害に役割を果たしており、CaM 活性化の数段階下流にもあります。
CaMKII は CaM によって直接活性化されるのに対し、PP1 は CaM のさらに数ステップ下流にあるという事実は、時間的ダイナミクスに大きな違いを生み出します。CaMKII はより迅速に活性化され、PP1 は活性化に時間がかかります。これらのダイナミクスは、基礎となる化学プロセスの詳細な測定を使用した以前のモデル ([[@BhallaIyengar99]]、[[@KurodaSchweighoferKawato01]]、[[@DoiKurodaMichikawaEtAl05]]) に基づいています。
{id=”figure_urakubo-poisson-25-100” style=”高さ:20em”}
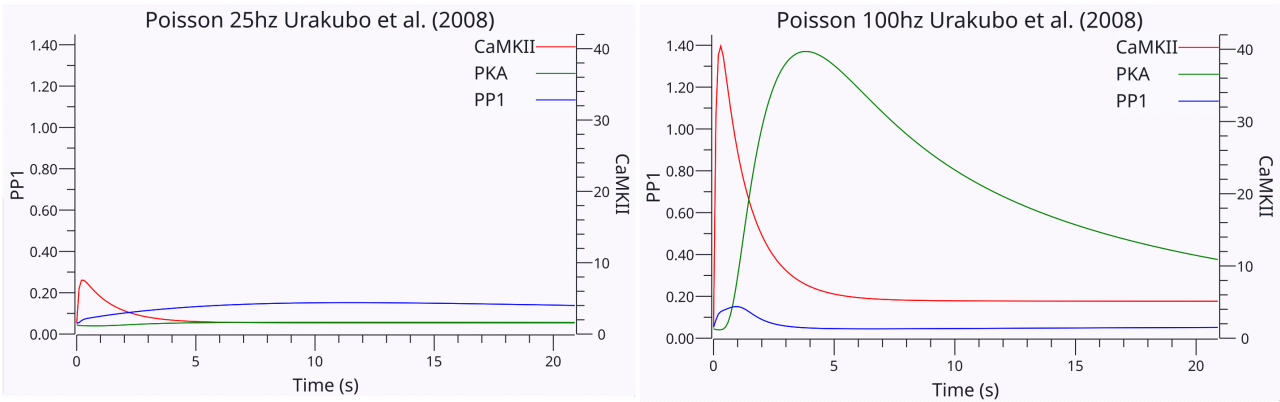
これらの化学プロセスがスパイク前後の活動に応答してどのように作用するかを確認するために、[[#figure_urakubo-poisson-25-100]] は、[[@^UrakuboHondaFroemkeEtAl08]] モデルの複製における 200 ミリ秒のポアソン ランダム スパイクに応答した CaMKII、PKA、および PP1 の時間経過を示します。最初の図の 25Hz ランダム活動のケースでは、PP1 の全体的な時間経過が長く、最終的には CaMKII の一過性活動を上回り、その結果 LTD (シナプス後密度からの AMPA 受容体のエンドサイトーシス、PSD) が発生することがわかります。 2 番目の図の 100hz ランダム活性のケースでは、CaMKII は最初は十分に活性化されて時間が経っても活性を維持し、高レベルのカルシウムと CaM 活性化による PKA 経路のより強力な活性化により、最終的に PP1 経路が阻害されます。これにより、LTP (シナプス後密度における AMPA 受容体の発現) が生じます。
{id=”figure_urakubo-xcal” style=”高さ:30em”}
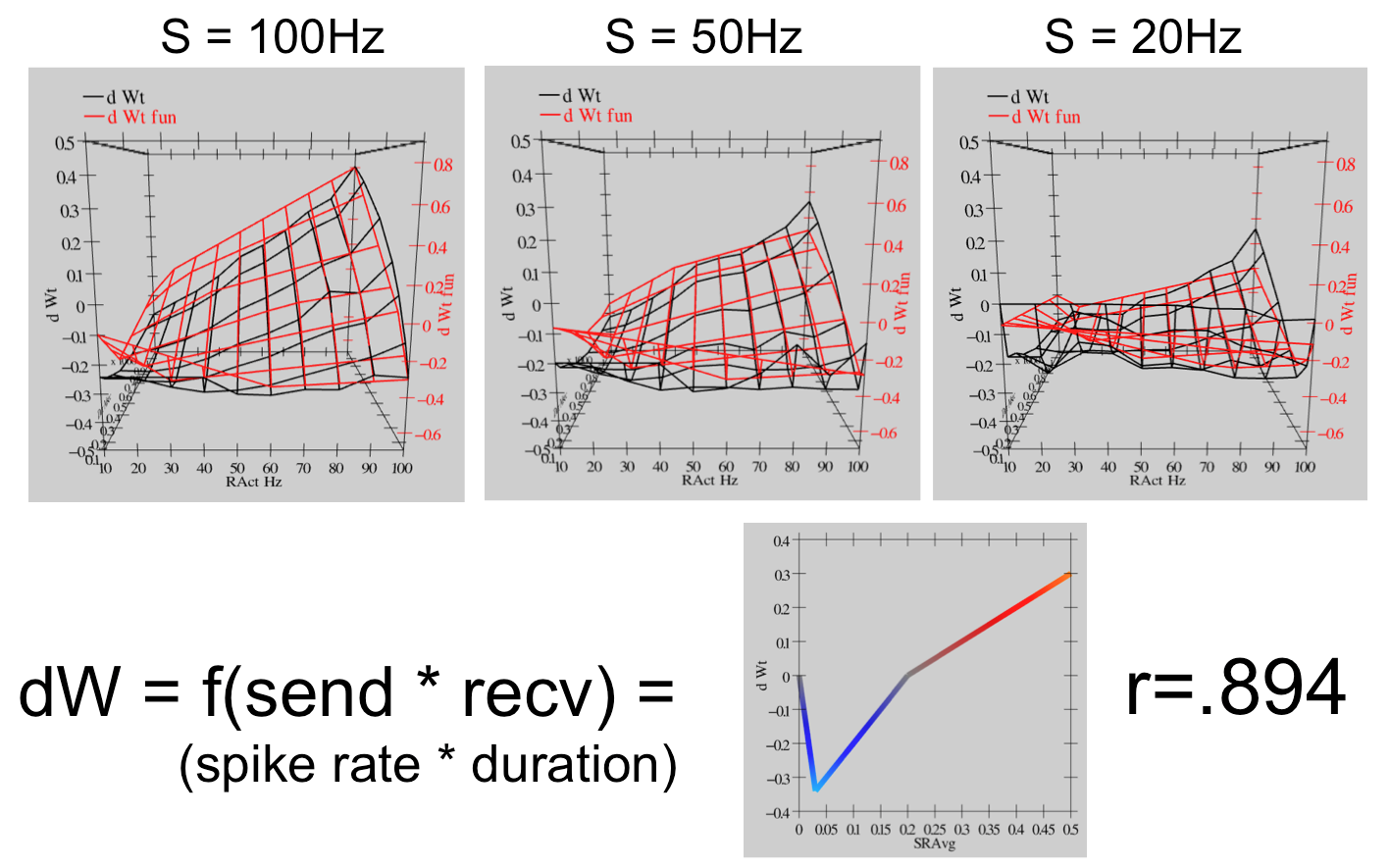
さらに、さまざまな周波数と持続時間のスパイク列を系統的にプローブすることにより、このモデルが [[#figure_ltpd-ca]] CaMKII 対 PP1 モデルによって予測される [[#figure_ltpd-ca]] に示される BCM のような曲線をどの程度再現するかをテストしました。 [[#figure_urakubo-xcal]] に示されている結果は、このモデルが広範囲のアクティビティ パラメーターにわたって BCM のような動作を示し、BCM 関数の単純な線形化バージョンが高レベルの分散 (r=0.894) を捕捉していることを裏付けています。このデータは [[urakubo08 simulation]] で自分で複製できます。
{id=”figure_urakubo-stdp” style=”高さ:20em”}
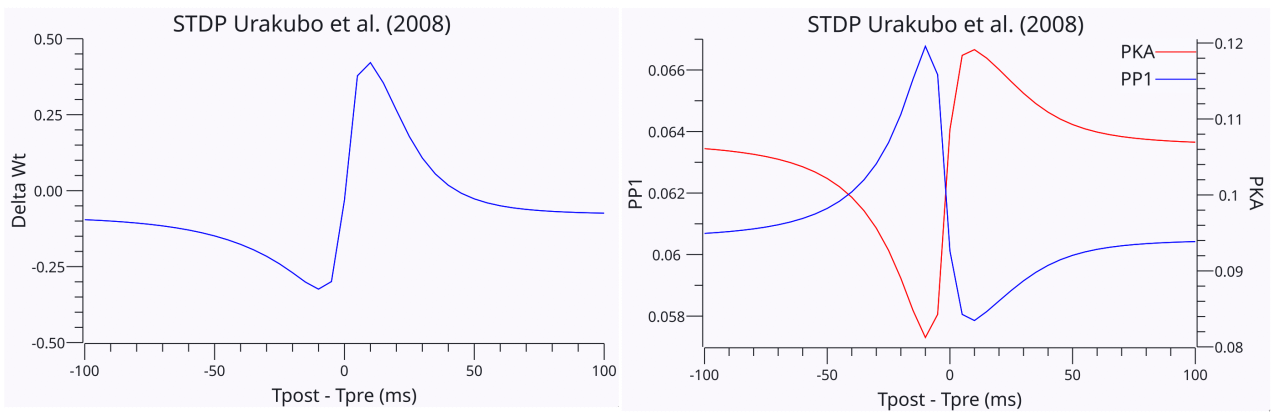
[[@^UrakuboHondaFroemkeEtAl08]] モデルは、[[#figure_urakubo-stdp]] に示すように [[@^BiPoo98]] STDP 結果を複製することもできますが、これは STDP 現象を直接サポートする非常に特殊な変更が必要な場合に限ります。具体的には、グルタミン酸が NMDA 受容体に結合すると、その不活化速度が 2.3 ms (非結合) から 230 ms (結合) へと 100 倍変化する「アロステリック」ダイナミクスを実装しました。アロステリックという用語は、反応に直接関与する部位とは異なる部位に結合することによるタンパク質機能の調節を指します (これは非常に一般的です)。したがって、ここではグルタミン酸が、カルシウムの侵入の結果として、CaM 結合によって駆動される不活化反応を調節しています。したがって、シナプス前ニューロンが最初に発火すると、NMDA受容体に結合するグルタミン酸により、かなり長く開いたままとなり、高いLTPレベルのカルシウムが流入することが可能になります。逆に、シナプス後がシナプス前よりも先に発火する場合、NMDA 受容体の不活化時定数ははるかに短くなり、LTD レベルのカルシウムのみが入ります。
この仮説メカニズムは追加のデータによって裏付けられていますが、NMDA チャネルの十分に確立された特性とみなせるかどうかは不明です (たとえば、NMDA チャネルの不活性化メカニズムに関する最近の徹底したレビューでは特に言及されていません; [[@IacobucciPopescu24]])。これは、[[@^UrakuboHondaFroemkeEtAl08]] モデルに欠けている他のメカニズムが、代わりに STDP 現象の背後にある関連する差別化要因を提供するのではないかという疑問を引き起こします。それにもかかわらず、STDP のようなパラダイムのより複雑なバージョンに関する [[@^UrakuboHondaFroemkeEtAl08]] モデルからの広範な追加結果でも、以前に議論したように、単純な因果関係の STDP ストーリーではシナプス可塑性の性質を適切に説明できないという十分な証拠があります。
NMDA GluN2B に結合する CaMKII の構造的役割
上記の CaMKII 機能の「古典的」モデルは、その中心的な前提と大きく矛盾する多くの発見によって疑問視されてきました。その代わりに、キナーゼが通常行うことの標準的な触媒モデル ([[@BayerGiese25]]) ではなく、構造的結合メカニズムに基づいて、CaMKII がどのように機能するかについての新しい理解が生まれつつあります。具体的には、CaMKII 上の結合部位 Thr286 は、NMDA 受容体の GluN2B 位置に結合し、この結合は、CaMKII が関与するその後のシナプス可塑性段階の推進に重要であると考えられます ([[@TullisBayer23]]; [[@ChenCaiZhouEtAl24]])。
これは構造的な説明である。なぜなら、本質的な原因段階はその触媒活性化ではなく、NMDA 受容体に結合した CaMKII の物理的位置であるからである。それにもかかわらず、古典的なモデルの他のプロセスの多くは依然として関連しています。例えば、CaMKIIを活性化するためのCa++活性化CaM結合の最初のステップ(Thr286で起こる)は、そもそもGluN2Bに結合するために必要であるため、カルシウムの侵入によって活性化されたCaMKIIのみがこの構造結合に関与します。
対照的に、CaMKII タンパク質上のこの同じ Thr286 結合部位は、自己触媒的リン酸化 ([[@Lisman94]]) を通じて記憶痕跡を維持する能力に重要であるという仮説が立てられました。具体的には、CaMKII はこの結合部位を介して結合した 12 個のタンパク質要素の複合体を形成することができ、これが一度起こると、CaMKII を無期限に亘ってキナーゼ活性化の安定した状態に保ち、それによって長期記憶維持シグナルを提供するという理論がありました。しかし、蓄積されたデータは、CaMKII が活性化された Ca++-CaM 分子が周囲に存在するときにのみ一時的に活性化することを強く示唆しているため、長期記憶シグナルとしては妥当ではないようです ([[@BayerGiese25]])。
したがって、新しい構造モデルは、この一時的に活性化された CaMKII が、より長期間持続する記憶カスケードを駆動する方法を提供します。構造的な GluN2B 結合には一時的な活性が必要です。さらに、この構造的結合の結果は、[[@^Lisman94]] が最初に提案したもの、つまり Thr286 部位で結合した複数の CaMKII 複合体の安定化とおそらく類似しています。新しいことは、この CaMKII が PSD 内の NMDA 受容体の周りに集中していることです。これにより、PSD での AMPA エキソサイトーシス、アクチン線維の再構成、そして最終的なタンパク質合成を引き起こす複数のさらなるイベントを物理的に引き起こすのに適した場所にそれらの受容体が置かれることになります。
構造的 LTD メカニズム: DAPK1 および Thr305/306
CaMKII 機能の GluN2B 構造結合機構の重要な特徴の 1 つは、LTD と LTP の間の切り替えがどのように機能するかを理解する全く異なる方法を提供することです。GluN2B への CaMKII の結合を妨げるものはすべて LTD を生じ、一方、それを促進するものはすべて LTP を生じるはずです。
CaMKII GluN2B 結合を妨げ、LTD を引き起こす重要なメカニズムの 1 つは、これと同じ結合部位 ([[@GoodellZaegelCoultrapEtAl17]]; [[@GoodellTullisBayer21]]; [[@CookBuonaratiCoultrapEtAl21]]; [[@TullisBayer23]]) への_DAPK1_ (death-associated protein kinase 1) の競合結合です。 DAPK1 は構造的および機能的に CaMKII ([[@BayerSchulman19]]) と類似していますが、より広範囲に発現しており、その名前が示すように、通常は細胞死 (アポトーシス) の誘発に関与しています。しかし、ニューロンでは、より特異的な機能があるようです。
{id=”figure_dapk1” style=”高さ:20em”}
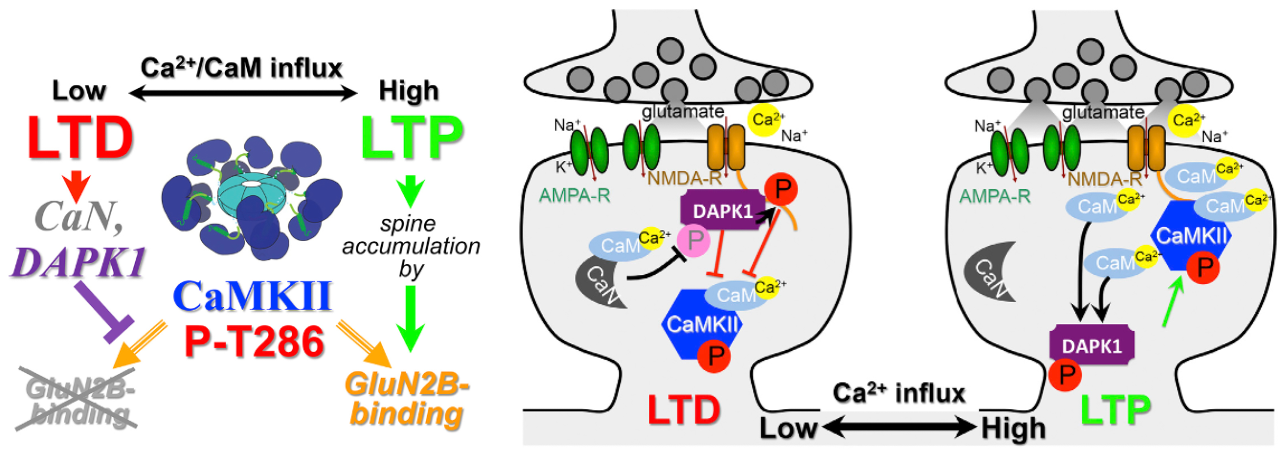
具体的には、[[#figure_dapk1]]に示すように、[[@^GoodellZaegelCoultrapEtAl17]]は、DAPK1上の異なる結合部位(S308)が活性化CaN(カルシニューリン)によって「脱リン酸化」された場合、DAPK1がGluN2Bに結合することを発見した(すなわち、これは、脱リン酸化が活性化をもたらすアロステリック機構である)。 [[#figure_urakubo-chem]] に示すように、CaN は Ca++ 活性化 CaM によって直接活性化されるため、DAPK1 は直接 Ca++ 流入から 1 段階離れていますが、CaMKII は CaM によって直接活性化されます。標準的な BCM のようなロジックを使用して、[[@^GoodellZaegelCoultrapEtAl17]] は、低レベルのカルシウムは GluN2B 部位への優先的な結合において CaN と DAPK1 の活性化に有利である一方、より高いレベルではこの 1 つの結合部位を巡る競合において CaMKII に有利であると主張しました。 CaMKII が勝った場合は LTP になりますが、DAPK1 が勝った場合は LTD となります。
{id=”figure_camkii-ltp-ltd” style=”高さ:15em”}
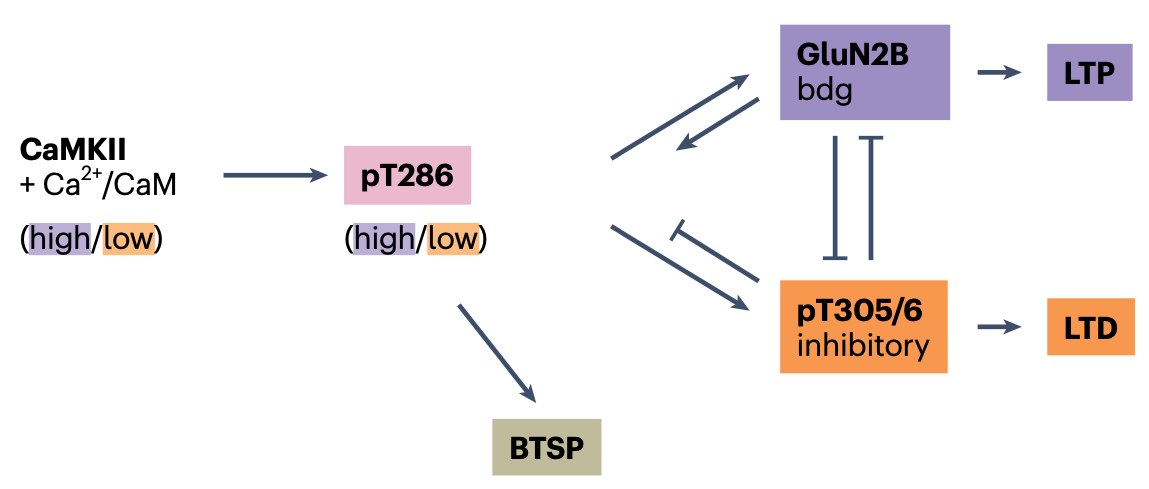
DAPK1 結合との競合に加えて、CaMKII タンパク質の異なる結合部位、Thr305/306 (Thr286 部位の近く) の活性化も、GluN2B 部位への CaMKII の結合を阻害します ([[@CookBuonaratiCoultrapEtAl21]]; [[#figure_camkii-ltp-ltd]])。この結果は、Thr305/306とDAPK1の活性化のいずれかを遮断するとLTDが阻止されるため、LTDを誘導するには両方の活性化が必要であることを示しています。 Thr305/306 部位の活性化は、他の CaMKII タンパク質による結合 (すなわち、自己触媒または自己リン酸化) を介して起こります。さらに事態をさらに複雑にしているのは、この自己触媒作用には Ca++-CaM による Thr286 の初期活性化が必要であり、これは LTP と LTD の両方に必須であることが以前に示されています ([[@CoultrapFreundOLearyEtAl14]])。
[[kinase algorithm]] の根底にある [[temporal derivative]] 機構に特に関連するのは、この Thr286 および Thr305/306 の自己リン酸化の時間的動態により、Ca++-CaM 結合による活性化の速いパターンと遅いパターンに対する強い感受性の差異が生じます。具体的には、[[@^CookBuonaratiCoultrapEtAl21]] で説明されているように、Thr286 での Ca++-CaM 活性化に対する素早い初期応答があり、その部位での複数の CaMKII 分子の結合を促進します。これが持続すると、その後 NMDA 上の GluN2B 部位に結合し、LTP が生じます。しかし、CaM 分子は Thr286 部位から比較的早く解離し、これが起こると、代わりに CaMKII 分子が近くの Thr305/306 部位で結合する傾向があり、これにより GluN2B 結合が阻害されます。
したがって、Thr286 対 Thr305/306 活性化の相対的なバランスは、シナプス活動の全体的な速度の違いを検出するのに理想的な位置にあると考えられます。特に、最初は強くなるが、その後低い速度に低下するシナプス活動のパターンは、Thr305/306 結合を活性化し、LTD を引き起こすのに理想的であると考えられます。これは、この活動パターンの LTD を示す [[Jang et al (2026)]] の結果と一致しており、計算的には予測誤差に相当します。つまり、最初の強い期待 (高いシナプス活動) が結果で確認されず、その後のシナプス活動の低下をもたらしました。
要約すると、DAPK1 とこの Thr305/306 の両方とも、LTP または LTD が発生するかどうかを決定するために互いに競合する、異なる実効時定数を持つ異なる経路の全体的な動態が、[[kinase algorithm]] が [[error-driven learning]] を実装するために使用する [[temporal derivative]] メカニズムの仮説メカニズムです。今後の研究の主な目標は、[[urakubo08 simulation]] を適応させて上記の GluN2B 構造メカニズムを組み込んで、その時間的応答プロファイルをより正確に決定することです。
より一般的には、シナプス可塑性に関するすべての研究から得られる重要な教訓は、誘導に使用される標準パラダイムが比較的限定されており、特に [[temporal derivative]] メカニズムに必要な種類の急速な時間的差異が含まれていないということです。したがって、[[Jang et al (2026)]] の結果が示すように、この研究は、非常に異なる周波数での定常状態の活動の単純なバースト、または高度に人為的な STDP パラダイムに焦点を当てることによって、LTP と LTD を駆動するための最も重要な条件を見落としている可能性があります。
神経調節および一時的適格性トレース
数 100 ミリ秒のタイムスケールでの急速な時間的ダイナミクスに加えて、シナプス可塑性の形成に重要と思われるより長いタイムスケールが存在します。たとえば、アクションが実行されてから、そのアクションに関連するその後の報酬または罰が経験されるまでの時間間隔を考えてみましょう。状況に応じて、この時間間隔は、1 秒から数時間、数日、さらにはそれ以上に及ぶ可能性があります。
アクションの実行に関連する最初のシナプス活動は、関連するシナプスの「適格性トレース」を開始し、その後の結果中に発生する活動によってシナプスが変更されることを可能にし、それによってこれら 2 つの時点にわたる重要な因果関係を確立すると考えられています。
[[hipppocampus]] の CA1 領域のニューロンからの最近のデータは、これらの「行動」タイムスケールの短い端に広がる学習偶発性の証拠を示しており、これは「行動タイムスケール シナプス可塑性」と呼ばれています (BTSP; [[@BittnerGrienbergerVaidyaEtAl15]]; [[@BittnerMilsteinGrienbergerEtAl17]]; [[@MilsteinLiBittnerEtAl21]]; [[@FanKimJenningsEtAl23]])。具体的には、これらのニューロンは、樹状突起の「プラトー電位」の上昇の結果として、空間調整の急速な変化を示すことがあり、関連するシナプス前の活動から数秒分離することができます。
BTSP におけるこれらのプラトー電位は、嗅内皮質からのシナプス入力の結果として発生し、目標の結果と関連しています。したがって、これらは、アクションの結果を示す一種の「第 3 要素」トレーニング信号 ([[@GerstnerLehmannLiakoniEtAl18]]) の一例であると考えられています。このような第 3 因子には、[[dopamine]]、[[acetylcholine]]、[[norepinepherine]]、[[serotonin]] などの神経調節物質が含まれます。これらはすべて、さまざまな方法で学習を調節することが示されており、報酬と罰の結果に関連しています。これらの神経調節物質のうちの 2 つ、[[norepinepherine]] および [[serotonin]] は、たとえば、皮質の可塑性に対して遅延 BTSP のような効果があることが判明しました ([[@HeHuertasHongEtAl15]])。
これらの現象の背後にある根本的なメカニズムの理解が得られるようになったのはつい最近ですが、最初の兆候は、CaMKII の中心的な役割やおそらく構造的な Thr286 結合 ([[@XiaoLiChitwoodEtAl23]]; [[@FanKimJenningsEtAl23]]; [[@JainNakahataPancaniEtAl24]]; [[@BayerGiese25]]) など、上で議論したのと同じメカニズムのすべてが関与しているということです。構造的に結合した CaMKII が永続的に存在することにより、CaMKII が関連する時間間隔に広がり、シナプス前入力イベントを後の結果ベースの信号と結び付けることが可能になる可能性があります。
これらの一時的な架橋メカニズムに関する別の初期の文献は、上記で述べた可塑性変化の_後期維持_に関与する明確なメカニズムに基づいて、1時間以上持続する可能性のある_シナプスタグ_に焦点を当てていました([[@MorrisFrey97]]; [[@ReymannFrey07]]; [[@RedondoMorris11]])。誘導事象をこれらの後のタンパク質合成依存機構と結び付ける必要性を考えると、この種の一時的な架橋はすでに誘導後の化学カスケードに必要な特徴であると思われる。
Axon で使用される [[kinase algorithm]] には、シナプス活動トレースを時間の経過とともに持続できるようにするパラメーターが含まれており、[[Rubicon]] フレームワークは、[[basal ganglia]]、[[amygdala]] およびその他の領域のそのようなトレース メカニズムに大きく依存して、行動に関連するタイムスケールにわたる接続を学習します。計算レベルでは、[[@^BellecScherrSubramoneyEtAl20]] は、同じ形式の適格性トレースを使用して、経時的な誤差逆伝播の形式をどのように達成できるかを示しました。