compcogneuro/web: thalamus
このページは外部資料の日本語訳です。原文の見出し順と本文順を保ち、コード・URL・出典表記はできるだけ原形のまま残しています。
出典とライセンス
原典: https://github.com/compcogneuro/web/blob/main/content/thalamus.md
ライセンス: Text: CC BY 4.0; code: BSD 3-Clause。このページは日本語翻訳であり、変更点は翻訳とサイト内整形です。
+++ Categories = [“Neuroscience”] bibfile = “ccnlab.json” +++ 視床は、複数の異なる脳回路において重要な役割を果たしているため、主にそれらの他の回路との関連で議論されます。このページでは、視床の組織的および機能的な全体像と、他のページへのポインターを提供します。視床の機能を理解することは、神経機能をより広範に理解するための強力な統合および橋渡しツールとなります。1 つのドメインの仕組みを理解すれば、同じ概念を他のドメインにも拡張できるからです。
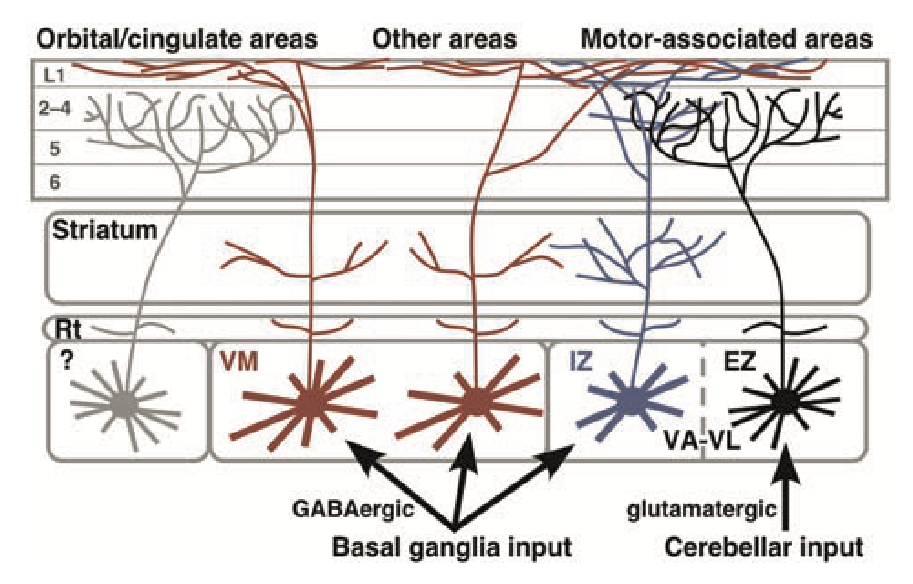
| [[evolution | evolutionary]] の比較の観点から見ると、視床は一次感覚野と、脳の最も外側の領域である パリウム、つまり テレセフェロン の間の相互接続を一貫して提供します。哺乳類では、[[neocortex]] はパリウムの主要な構成要素ですが、他の脊椎動物は、パリウム内に新皮質 ([[@Butler08]]) と同じ機能的役割を果たす、より小さく単純な皮質様構造を持っています。以下の機能は哺乳類に特有です。 |
-
注意を調節して一次感覚入力と運動入力を視覚のための LGN (外側膝状核) などの新皮質に中継します。感覚皮質領域からこれらの一次感覚視床領域に戻る広範なトップダウン投影があり、これらの感覚入力の注意調節をサポートしています ([[@PoltoratskiMaierNewtonEtAl19]]; [[@Briggs20]])。一次運動野は、[[cerebellum]] からの直接的な一次運動入力と、運動側で同様の役割を果たす体性感覚視床からの入力の両方を受け取り、[[motor#frontal cortex motor cortex]] に「脊髄の言語で話す」方法を教えます。 -
後部 [[neocortex]] では、視床の_髄核_がトップダウンの予測とボトムアップの結果を交互に表すことによって [[predictive learning]] をサポートします。同様の接続性は、[[prefrontal cortex]] および下部 [[motor]] 皮質領域と相互接続する_中背核 (MD)_ 核内のニューロンのサブセットにも存在し、同様の予測学習の役割を示唆しています。同様に、[[space]] を処理する脳システムに接続されている前視床領域は、この形式の学習をサポートできます。
-
AM、MD、VA / VM を含む複数の異なる視床核があり、[[basal ganglia]] (BG) からの脱抑制性出力信号を受信し、[[motor]] および [[prefrontal cortex]] (PFC) のすべての領域に集合的に投射し、そこで活動状態の急速な位相変化を引き起こすことができます (すなわち、gating。 [[PBWM]] モデル)、モーターの計画から実際の実行への移行中に明らかになるものなど。 [[Rubicon]] モデルでキャプチャされているように、PFC の中核となるゴール駆動領域では、これらのゲート信号がゴール選択からゴール関与状態への移行を駆動します。これらの前頭野には、
ITレイヤー 5 ニューロン ([[@ShepherdYamawaki21]]) として知られる、予測学習と BG ゲーティングを統合する予測学習ニューロンの重要なサブセットもあります。 -
層内 (IL) 核は、大脳基底核に対する広範なフィードバック経路を提供します。
- 室傍視床核は、目標駆動型 PFC、海馬、および関連する [[emotion]] 処理皮質下領域と相互接続し、目標駆動型処理の全体的な調整を提供します。
視床回路
{id=”figure_pulv-conns” style=”高さ:15em”}
![一次視覚野と二次視覚野の場合の、新皮質と視床の肺核の間の接続。網膜からの視覚入力は、視床の外側膝状核 (LGN) にドライバー入力を提供し、その後グルタミン酸作動性興奮性投射を主に一次視覚野の第 4 層 (V1) に送ります。髄様視床核は、V1 の深層 5b ニューロンから強力なドライバー入力を受け取り、視床中継ニューロン (TRN) ごとにドライバー シナプスが 1 つしかない可能性があります。二次視覚野 (V2) には、第 6 層 (VI) ニューロンからの比較的弱い投射が多数存在します。 [[@^ShermanGuillery06]] に基づく図。](https://raw.githubusercontent.com/compcogneuro/web/main/content/media/fig_pulvinar_connectivity.png)
[[#figure_pulv-conns]] は、[[@ShermanGuillery06]] に基づいており、哺乳類の脳の一次視覚野と二次視覚野の視床接続を示しています。この接続の主な特徴は、ほとんどの視床領域に共通しています。
| さまざまな視床核には、視床ニューロンの主要なタイプが 1 つだけ存在し、それが「視床中継ニューロン」 (TRN) です。視床本体を包み込む外殻である「視床網状核」 (TRN) は唯一の例外であり、以下で説明する GABA 作動性 [[inhibition | inhibitory]] ニューロンを持っています。 TRN は興奮性グルタミン酸神経伝達物質を [[neocortex]] に放出し、質的に異なる 2 種類の入力を受け取ります。 |
-
ドライバーは強力な入力であり、多くの場合、その数は比較的少ない (単一入力程度)。それらは、複数のシナプス接続を備えた大きく拡張された糸球体、さらには単一の解剖学的構造にカプセル化された「専用の」抑制性内ニューロンを持つことができます。これらは、皮質下領域(例えば、視覚のためのLGNへの網膜入力)または皮質領域(例えば、膝蓋骨によって実行される[[predictive learning]]のドライバーである深層5b IB固有のバーストニューロン)に由来する可能性があります。一部の視床領域にはドライバー入力が特定されておらず、通常は大脳基底核出力経路から広範な抑制性入力を受け取る領域があります。
The nature of the driver inputs is one of the most important clues to thalamic (and cortical) function. For example, cortical drivers from intrinsic bursting neurons provide the temporal dynamics needed to support predictive learning via the [[temporal derivative]] mechanism, whereas subcortical sensory drivers do not appear to have this property. However, subcortical drivers in the motor system naturally have the requisite delays, as discussed in the [[motor#Frontal cortex learning via subcortical efferent copy signals motor]] page. -
非ドライバーは残りの入力であり、脳内の他のほとんどのシナプス接続と同様です。 [[@ShermanGuillery06]] や他の多くの研究者は、これらを「変調入力」としてラベル付けしていますが、[[predictive learning]] のコンテキストで提案されている関数を考えると、これは機能的な仮定が強すぎる可能性があります。たとえば、[[@OReillyRussinZolfagharEtAl21]] でレビューされているように、これらの入力が TRN の持続的な発射を駆動できるという十分な証拠があります。
TRN は特に局所的な興奮性相互接続性を欠いており、TRN 網様ニューロンによって媒介されるプールされた抑制の形態のみを持っています。したがって、それらは、受信した入力を忠実に表現するのに独特に適しており、それ自体に大きな歪みを与えない一種の「投影スクリーン」として機能します。
視床回路の比較的不活性な性質により、その回路はリレー システムとして特徴づけられていますが、2 つの特徴的なタイプの入力の存在は、より強力な種類の機能がサポートされている可能性があることを示唆しています。たとえば、[[predictive learning]] フレームワークは、粉体へのドライバー入力の特有の時間的側面が、時間にわたるドライバー入力と非ドライバー入力間の誤差勾配を効果的に計算する [[temporal derivative]] 計算をどのようにサポートするかを示しています。
TRN から皮質への接続の性質は、視床の機能についての別の重要な洞察を提供します。これらの投影が変化する主な次元が 2 つあります。これらはもともと [[@^Jones98]] の コア 対 マトリックス カテゴリによって捕捉されましたが、より正確には次元 ([[@ClascaRubio-GarridoJabaudon12]]; [[@ShermanUsrey24]]) として分類されます。
-
層流ターゲット: 古典的な core タイプは主に皮質の中間層、主に層 4 (例: [[#figure_pulv-conns]] に示すように) に投影しますが、古典的な matrix タイプは層 1 と 5 に投影しますが、中間層は避けます。ただし、実際には、明確に定義されたカテゴリーではなく、異なる視床ニューロン間でこれらの接続の割合が異なります。
Functionally, projections into the middle layers provides direct input information into the cortex, as in the prototypical sensory input into primary sensory areas. Projections into layers 1 and 5 can drive two main functions: regulation of overall cortical activity levels via the VIP inhibitory interneurons in layer 1 (this was the original interpretation of the matrix type), and maintenance of sustained activity (primarily in [[prefrontal cortex]]) via bidirectional excitatory loops through layer 5b PT (pyramidal tract) neurons, which also have extensive dendritic tufts in layer 1.
-
領域焦点: 古典的な core タイプは、通常、比較的限定された focal 方式で 1 つの皮質領域のみに投影しますが、古典的な matrix タイプは、より広範囲に、複数の異なる領域に拡散して、より調整的な機能と一致して投影します。
{id=”figure_map-phillips” style=”高さ:30em”}

[[#figure_map-phillips]] は、げっ歯類のさまざまな視床領域のマップを示しています。これは、遺伝子発現のパターンに基づいて 3+1 の幅広いカテゴリのセットを提供しており、さまざまな視床領域にわたる機能の理解を知らせるのに役立ちます。このデータは、接続パターンに関してより詳細な区別を提供しませんが、以下で説明するように、一般にさまざまなタイプのドライバー入力に対応します。
{id=”figure_types-clasca” style=”高さ:30em”}

[[#figure_types-clasca]] は、[[@^ClascaRubio-GarridoJabaudon12]] による古典的なコアおよびマトリックスのカテゴリの拡張を示します。2 つの異なる M タイプがあり、そのうちの 1 つは古典的なマトリックス タイプに対応し、M タイプ (フォーカル) はコアとマトリックスの機能を組み合わせています。また、層内 (IL) 核の独特な投影パターンもあります。これには、大脳基底核の機能に重要な役割を果たし ([[basal ganglia#Parafascicular feedback loops]] を参照)、深層皮質層への投影とともに線条体への広範な接続性を持つ PF (筋膜傍) 核と CM (セントロメディアン) 核が含まれます。
コア対マトリックスの分類的アプローチを維持する代わりに、さまざまな視床領域のニューロンの接続プロファイルを説明する際に、機能的で特徴ベースの用語を使用することを好みます。特定の TRN セルは通常、これらの特性を複数持ちます。
-
入力: 中間層への焦点投影により、皮質領域への情報入力を推進します。
-
メンテナンス: レイヤー 1 および 5 への焦点投影。レイヤー 5b PT ニューロンのアクティブなメンテナンスをサポートします。
-
調節性: 層 1 および 5 への拡散投影。これは古典的な覚醒調節マトリックス型ニューロンに限定されるものではありません。VA および VM を含む大脳基底核調節運動野にはこのパターンがあり、運動皮質領域のより動的で細かい調整を提供します。
-
単一: 単一のエリアに投影します。
-
マルチ: 複数のエリアにプロジェクトを投影します。これは、投影の焦点性と拡散性の性質に直交する可能性があることに注意してください。たとえば、MDm (内側背側) には焦点性がありながら複数領域の投影があります。
視床の機能組織
{id=”figure_map-clasca” style=”高さ:30em”}
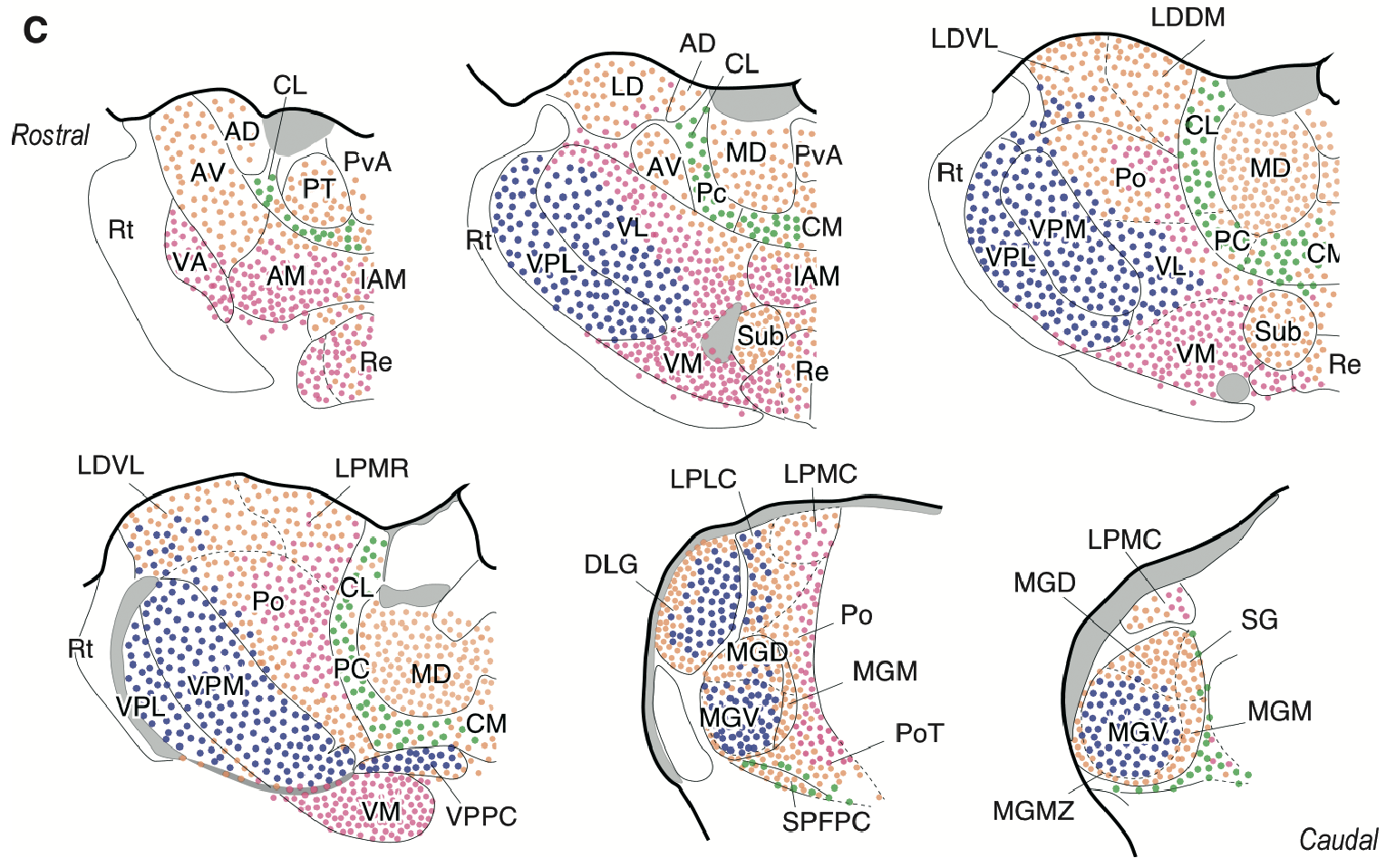
{id=”figure_map-rovo” style=”高さ:50em”}
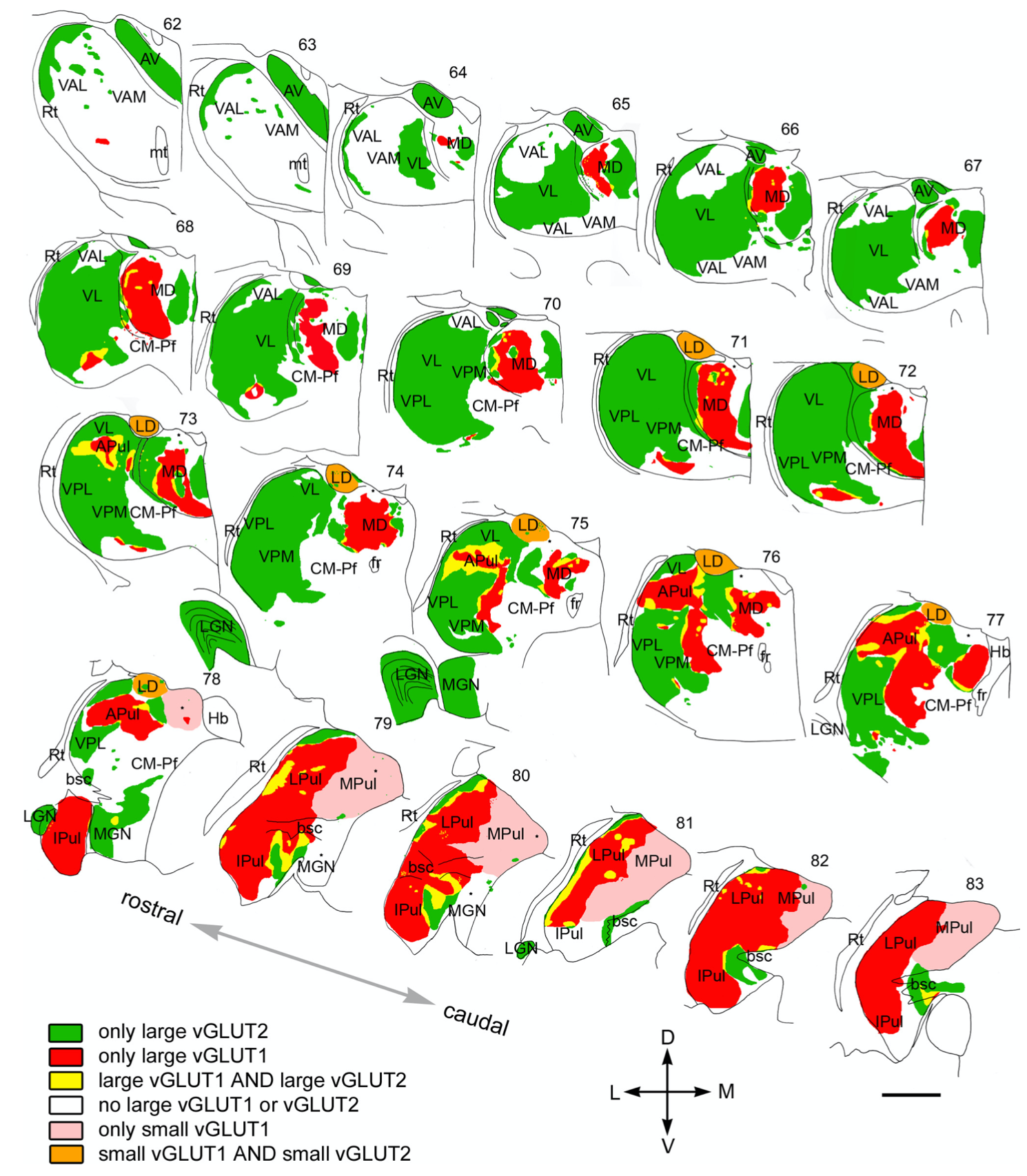
[[#figure_map-clasca]] は、[[@^ClascaRubio-GarridoJabaudon12]] カテゴリを使用したげっ歯類の視床のマップを提供し、[[#figure_map-rovo]] は、各領域に存在するドライバー シナプスのタイプをラベル付けしたマカクザルの視床のマップを提供します ([[@RovoUlbertAcsady12]])。これらは機能的に次のように解釈できます。
- 皮質下ドライバー: 強力 = 大きい vGLUT2 (緑色)。
- 皮質 ドライバー: 強い = 大きい vGLUT1 (赤)、弱い = 小さい vGLUT1 (ピンク)。
- なし (白)。これらは通常、大脳基底核から抑制性入力を受け取ります。
- 他の組み合わせについては、該当する場合に以下で説明します。
これら 2 つのマップを組み合わせることで (そして [[#figure_map-phillips]] のデータを念頭に置くと)、さまざまな視床領域の一般的な機能を特定できます。
各視床核には、私たちがそれを識別する主要なタイプの TRN 接続パターンがありますが、ほとんどの場合、ある程度の多様性が存在することを認識することが重要です。たとえば、古典的なマトリックス タイプのニューロンが、LGN ([[@ShermanUsrey24]]) で優勢な古典的なコア タイプの中に混在しているなどです。機能的には、これらのマトリックス型ニューロンは幅広い接続パターンで多くの領域をカバーできるため、仕事を行うために必要なニューロンは少なくなります。一般に、そのようなニューロンを介した皮質の全体的な興奮性と覚醒レベルの調節は、ほとんどの視床領域によって実行される追加の仕事であると想定されますが、この機能のみを実行する専用の領域は不要です。
-
一次感覚 (ドライバー: 皮質下; 投影: 入力、単一): LGN (外側膝状核、視覚、V1); MGNv (内側腹側 GN; 聴覚、A1); MGNm (内側内側 GN、前庭など); VPL、VPM (腹側後外側、外側、内側、体性感覚、S1)。これらは、VL と AV を除き、ドライバー図のほとんどの緑色の領域であり、皮質下ソース (LGN の場合は網膜など) からの大きくて強力なグルタミン酸作動性ドライバーがあります。
-
一次運動 (ドライバー: 皮質下、投影: 入力、単一): VL (腹側外側)。これは、一次タイプの視床領域であり、感覚ではなく、一次 [[motor]] (M1) です。これらのニューロンに対する強力な興奮性皮質下ドライバーは、[[cerebellum cerebellar]] 核 (CN) に収束し、そこから VL に投射する上行性脊髄経路から発生します。これらのニューロンは、脊髄運動システムに送られたコマンドに対して実際に行ったことを反映する遠心性コピー信号など、一次運動野に複数のトレーニング信号を提供します。解剖学的には、VL は体性感覚 VPL、VPM 核に隣接しており、M1 から中央溝を挟んで地形的に一次体性感覚皮質 (S1) に投影されています。 [[#frontal thalamus]] 領域の詳細については、以下を参照してください。 -
一次空間と時間 (ドライバー: 皮質下; 投影: 入力、複数): AV (前腹側) および AD (前背側)。 VL と同様に、これは皮質下で駆動され、標準的な感覚入力ではない入力投影領域の興味深いケースです。これらの領域は、乳頭体 (AV の内側と AD の外側) からドライバー入力を受け取り、重要な [[space spatial]] 信号と [[time timing]] 信号を伝え、その後、皮質の主要な空間表現領域である (前後) 海馬台および後脾皮質に投影されます。 AD 核に突き出ている側乳頭体は、頭方向細胞の主な供給源であり、前庭信号と運動信号を統合して、頭 (およびより一般的には身体) が (世界ベースの) 座標系でどの方向を向いているかを追跡します。房室に突き出た内側乳頭体は、腹側被蓋核 (VTN) で生成された [[theta rhythm]] 信号を伝えます。この信号は内側中隔のニューロンも駆動し、内側側頭葉と [[hippocampus]] 全体の処理を調整します。
-
二次知覚 (ドライバー: 皮質; 投影: 入力、変調、シングルおよびマルチ): 枢軸 はこのタイプの主要な核であり、すべての高次の感覚野への地形的投影を伴います。ドライバーマップはこれらの領域を赤とピンクで示しており、赤い領域には強力なグルタミン酸作動性ドライバーがありますが、一次領域とは異なり、これらは皮質下ドライバーではなく、層5bの皮質ドライバーに由来しています。齧歯動物では歯槽骨自体はそのように標識されていないが、外側後部 (LP) は接続性 ([[@LeowZhouSullivanEtAl22]]) に基づいて相同な領域であると考えられており、同じ焦点接続性 (LPMC、LPMR) を持っています。ドライバー マップの黄色の領域は、皮質と皮質下の両方の入力、特に肺骨内に突き出ることが知られている上丘からの入力を受け取る肺骨の領域を表します。
-
セカンダリ モーター (ドライバー: なし; 投影: 変調、メンテナンス、入力、マルチおよびシングル): VA / VM、AM (VAM、VAL [[#figure_map-rovo]] を含む前腹部、内側および前内側) は、強力なドライバーがないため、ドライバー図ではすべて白で色付けされています。代わりに、大脳基底核出力核 (SNr、GPi) からの広範な抑制とともに、さまざまな皮質下ソースからの収束入力を受け取ります。これらは接続図の赤い点の領域すべてであり、M 型多領域ニューロンの主要集団を表しています。この接続パターンに基づくと、これらは対応する高次運動野に非常に広範な調節効果を及ぼす可能性が高く、BGがBG出力経路の脱抑制ゲートを介してこれらの運動野における全体的な興奮性を調節できるようになります。詳細については、以下の [[#frontal thalamus]] を参照してください。
-
セカンダリ PFC、ゴール ゲート (ドライバー: なし; プロジェクション: 入力、メンテナンス、変調、マルチ): MD (背側)。げっ歯類では、MD のさまざまな領域 (内側、中央、外側) がさまざまな BG 脱抑制ゲート信号を受信し、複数の領域でありながら焦点を合わせて PFC のすべての目標駆動領域の中層に投射し ([[@KuramotoPanFurutaEtAl17]]; [[@PhillipsKambiRedinbaughEtAl21]])、複数のレベルの「ゴールゲート」をサポートします。 MDm 領域は、腹側淡蒼球 (VP) ([[#figure_map-rovo]] では白で表示) から広範な投影を受け取り、主要なゴール ゲーティング領域を表すすべての目標主導型 PFC 領域に投影します。 MDc と MDl (および霊長類の関連領域) はより多様な接続性を持っており、次に説明します。
-
二次 PFC、予測学習および目標更新 (ドライバー: 皮質; 投影: 入力、維持、単一): MDc、MDl (背背中央、外側)。 MD のこれらの領域内のニューロンのサブセットは、M 型の焦点接続パターンを持ちながら、強力な皮質ドライバー入力 ([[#figure_map-rovo]] の赤) を受け取るという点で、前頭前野の枢軸に実質的に似ています。予測学習をサポートするプルビナの強力な理論的および経験的根拠に基づいて、これと同じ機能が、これらの MD 領域が投影する OFC、IL、PL、および ACC PFC の目標主導型領域でもサポートされる可能性があります。これらの MD 領域の他のニューロンは、GPi および SNr から広範な脱抑制性 BG 入力を受け取り、それぞれが目標値システムの 2 つの別個のコンポーネントに投影します。OFC、IL = 結果値 (MDc)。 ACC、PL、ALM / dlPFC = アクションの価値 / コスト (MD1)。
-
層内 (IL: PF、CM など) (ドライバー: なし; 投影: 線条体、皮質深層への IL タイプ)。これらの核は、[[basal ganglia]] および運動新皮質の深層に広範なフィードバックを提供し、BG の機能と学習において重要な特殊な役割を果たします。
[[@GiguereGoldman-Rakic88]] は、霊長類における斑状の組織を示す霊長類の研究です。
前頭視床
前頭領域と相互接続されている視床領域には 3 つの主要な機能があります。
-
プライマリ [[motor#frontal cortex motor cortex]] が脊髄言語 (VL、POm / PULm) を話すためのトレーニング。これらは、脊髄運動信号 (VL) または体性感覚信号 (POm / PULm) からの強力なドライバー入力を受け取り、強力なドライバー入力を運動野の中層に投影します。 -
皮質皮質 [[predictive learning]]。後皮質の肺床によってサポートされるものと類似しているため、上位層は下位層の運動関連活動を予測する方法を学習できます。これは、MD 核および AM 核内のニューロンのサブセットによってサポートされています。これは、たとえば、より長い一連の運動動作の学習に役立ちます。
- 複数のタイムスケールにわたる前頭部表現の [[basal ganglia]] (BG) ベースの迅速な更新 (gating) の 2 つの形式: 目標達成のための活性化ベースの作業記憶更新 ([[Rubicon]] モデルによる) と、[[PBWM]] (前頭前野、大脳基底核の作業記憶) モデルと一致する運動計画の更新。もう 1 つは、運動動作の開始や計画の更新に関連する、より拡散した広範な同期形式のゲートです。作業記憶ゲート機能は、BG 脱抑制出力を受け取る MD および AM 内のニューロンのサブセットによってサポートされており、広範な同期機能は、霊長類に特有の特別な VAmc 核を含む VA (別名げっ歯類の VM、[[@IlinskyKultas-Ilinsky87]]) 核によってサポートされています。
げっ歯類
{id=”figure_frontal-alonso” style=”高さ:45em”}
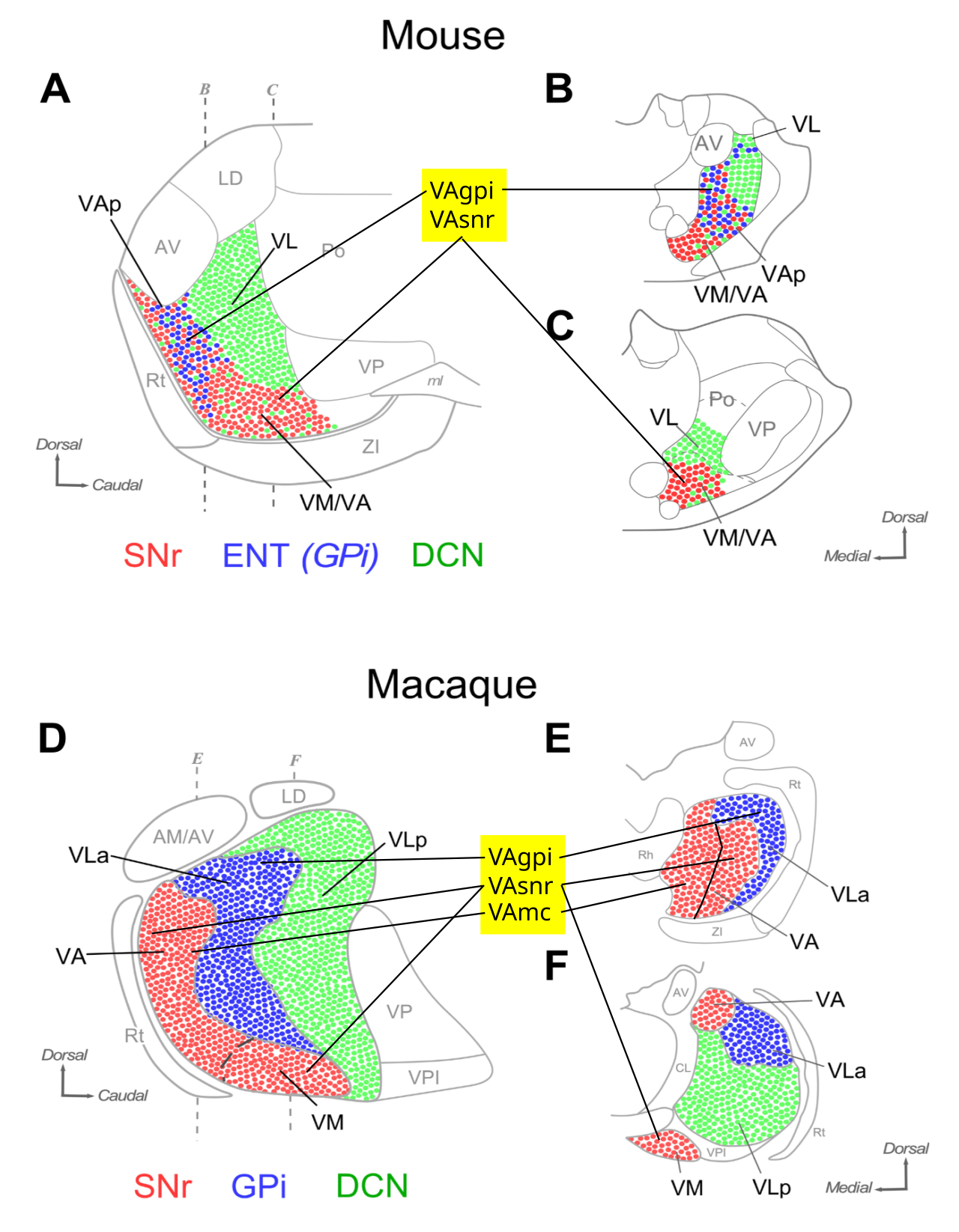
齧歯動物のより単純なケースから始めます。齧歯動物は、これらの異なる機能のそれぞれが異なる視床核によって明確に表されています。霊長類の場合は、これと同じ基本基盤の上に構築されていると理解できるさらに複雑さを追加します。 [[#figure_frontal-alonso]] は、前頭葉皮質 ([[@Alonso-MartinezRubio-TevesCasas-TorremochaEtAl23]]) に投射する腹側前視床核に対する BG (SNr、GPi) 対小脳 (CN) ドライバー入力の構成を示します。これらの視床領域の解剖学的用語は論文間および種間で一貫していないため、以降の議論で使用する次のラベルを定義します (同様の組織については [[@SieveritzKiani24]] を参照)。
VAbgは、BG 出力核から抑制性入力を受け取り、運動皮質には投射しますが、PFC 野には投射しない VM/VA/VL 野であり、具体的には次のとおりです。VAgpiは GPi (げっ歯類の脚内腔または ENT) から抑制性入力を受け取ります。VAsnrは SNr から抑制入力を受け取ります。げっ歯類では、これには吻腹側 VA ([[@Alonso-MartinezRubio-TevesCasas-TorremochaEtAl23]]) が含まれます。
VAmcは、大細胞を有する霊長類の VA のサブセットであり、PFC に投射し、SNr から受け取ります (げっ歯類には存在しません)。 ※VLcnはCN(小脳核)からの興奮性ドライバー入力を受け取り、一次運動(M1)に投射するVL野です。げっ歯類では、これには尾背側 VA ([[@Alonso-MartinezRubio-TevesCasas-TorremochaEtAl23]]) も含まれます。
{id=”figure_prjn-倉本” style=”高さ:25em”}
[[#figure_prjn-kuramoto]] ([[@KuramotoOhnoFurutaEtAl15]] より) は、齧歯動物の腹側視床経路を示しており、上記の標準ラベルに従って、ドライバー入力に従って識別できます。 VLcn は小脳核から強力なドライバーを受け取り、運動野の中層(つまりコアのようなもの)に焦点を当てて投射し、運動野が脊髄の言語を学習できるようにします。この投影は、霊長類の M2、SMA、および運動前野、およびげっ歯類の対応する高次運動野に当てはまります ([[@IlinskyKultas-Ilinsky87]])。
VAbg は BG 脱抑制出力 (VAsnr = SNr、VAgpi = GPi) を受け取り、広範囲に分岐した方法 (つまりマトリックス状) で表層に投射します。これにより、BG は広範なゲート信号を運動皮質全体に、さらにはそれを超えて送信することができ、一部のニューロンは後部皮質の広範囲の領域にも投射します。さらに、VA-VL の抑制ゾーン内にあるこれらのニューロンのサブセットが、OFC および ACC 領域に有利なより制限された接続を持っていることを示す兆候がありました ([[@KuramotoOhnoFurutaEtAl15]])。詳細については次のセクションを参照してください。しかし、全体として、これらのニューロンは、運動の準備から運動の実行までの移行など、重要な移行の時点ですべての運動野を同期させるのに重要であると考えられます。見る
{id=”figure_md-pfc” style=”高さ:20em”}
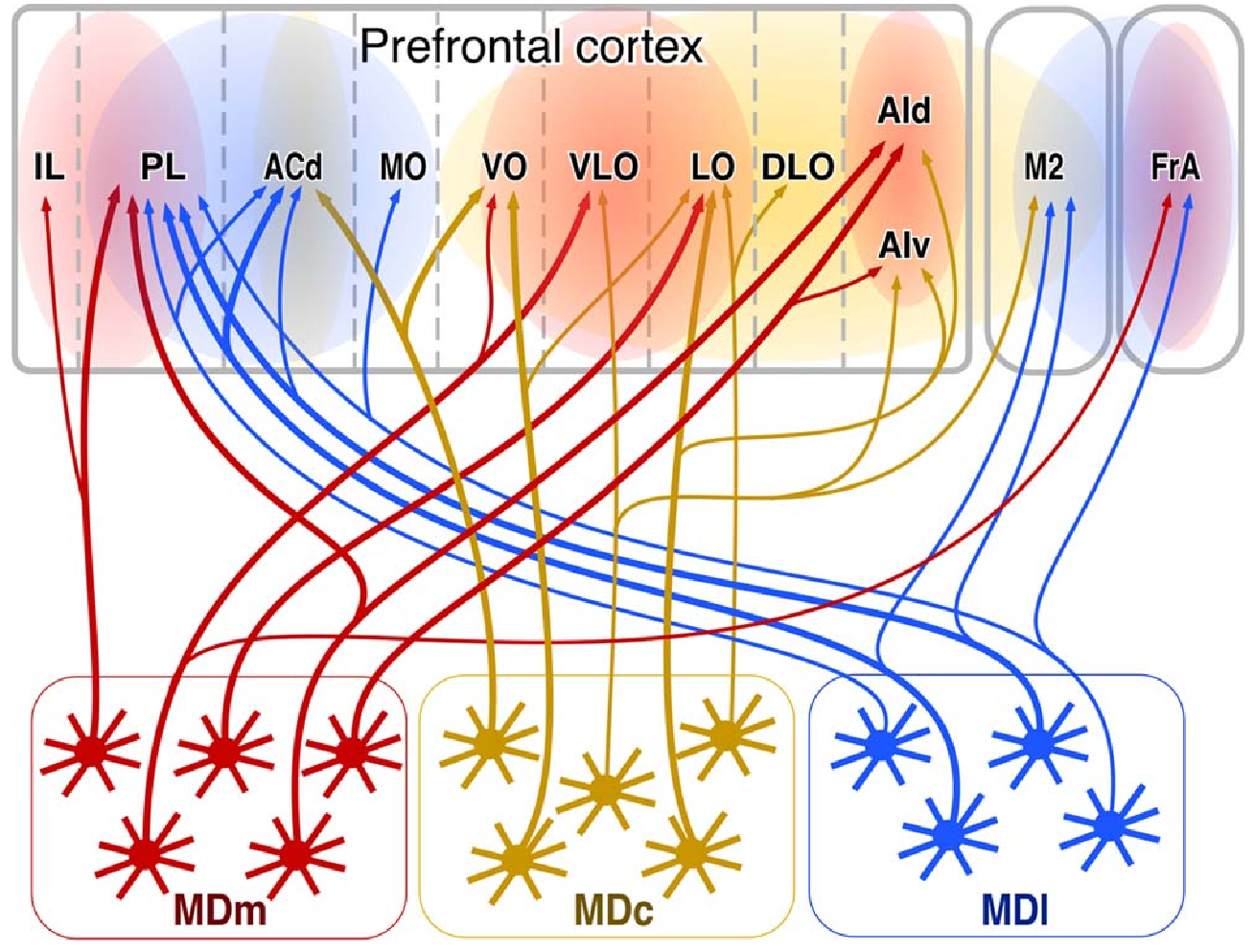
[[#figure_md-pfc]] は、ラットの PFC 領域と相互接続する MD 視床のさまざまな領域 ([[@KuramotoPanFurutaEtAl17]] より) を示します。MDm (内側) によってサポートされる主要なゴール ゲーティング機能は、すべてのゴール領域 (IL、PL、OFC、および FrA (ALM)) に投影され、出力である VP (腹側淡蒼球) からの脱抑制を受け取ります。 [[basal ganglia]] に記載されている腹側線条体ゴール ゲート領域からの経路。 MD の他の 2 つの領域 (c = 中央、l = 側方) は、OFC (中央) と運動、ACC、および PL (側方) に相補的でより特異的なゲート経路を提供し、潜在的に、目標表現のこれらの調整されたコンポーネントのそれぞれを個別に更新できるようにします。 MD 投影は、レイヤー 1 と深層 ([[@CollinsAnastasiadesMarlinEtAl18]]) をターゲットとする VA (VM) からの投影と比較して、中間層 (2、3) への方がはるかに強力です。
これらの MD 領域には、おそらく CT 入力 (第 6 層から) と皮質ドライバー (第 5 層の PT ニューロンから) を受け取り、[[predictive learning]] ([[@CollinsAnastasiadesMarlinEtAl18]]) の肺門と同様に機能する非 BG ゲート ニューロンもあります。 [[Rubicon]] モデルは、たとえば、より抽象的な IL 値ニューロンが OFC 固有の結果表現を予測するのに対し、PL は IL と ACC の両方の活動を予測するようにこれらを組織化します。 ACC は皮質下の行動コスト信号を予測することで学習し、OFC は BLA ([[amygdala]]) 結果信号を予測することによって学習します。
げっ歯類の後内側 (POm) 核は、一次感覚皮質 (S1) からの体性感覚信号の高次視床領域であり、視覚およびその他の高次感覚処理領域に関連して肺床への接続性が類似しています ([[@MoPetrofViaeneEtAl17]]; [[@LandismanConnors07]])。たとえば、一次体性感覚視床核(VPM)と比較して、POm のみが第 5b 層の PT ニューロンからドライバー入力を受け取ります。
霊長類
{id=”figure_frontal-areas” style=”高さ:40em”}
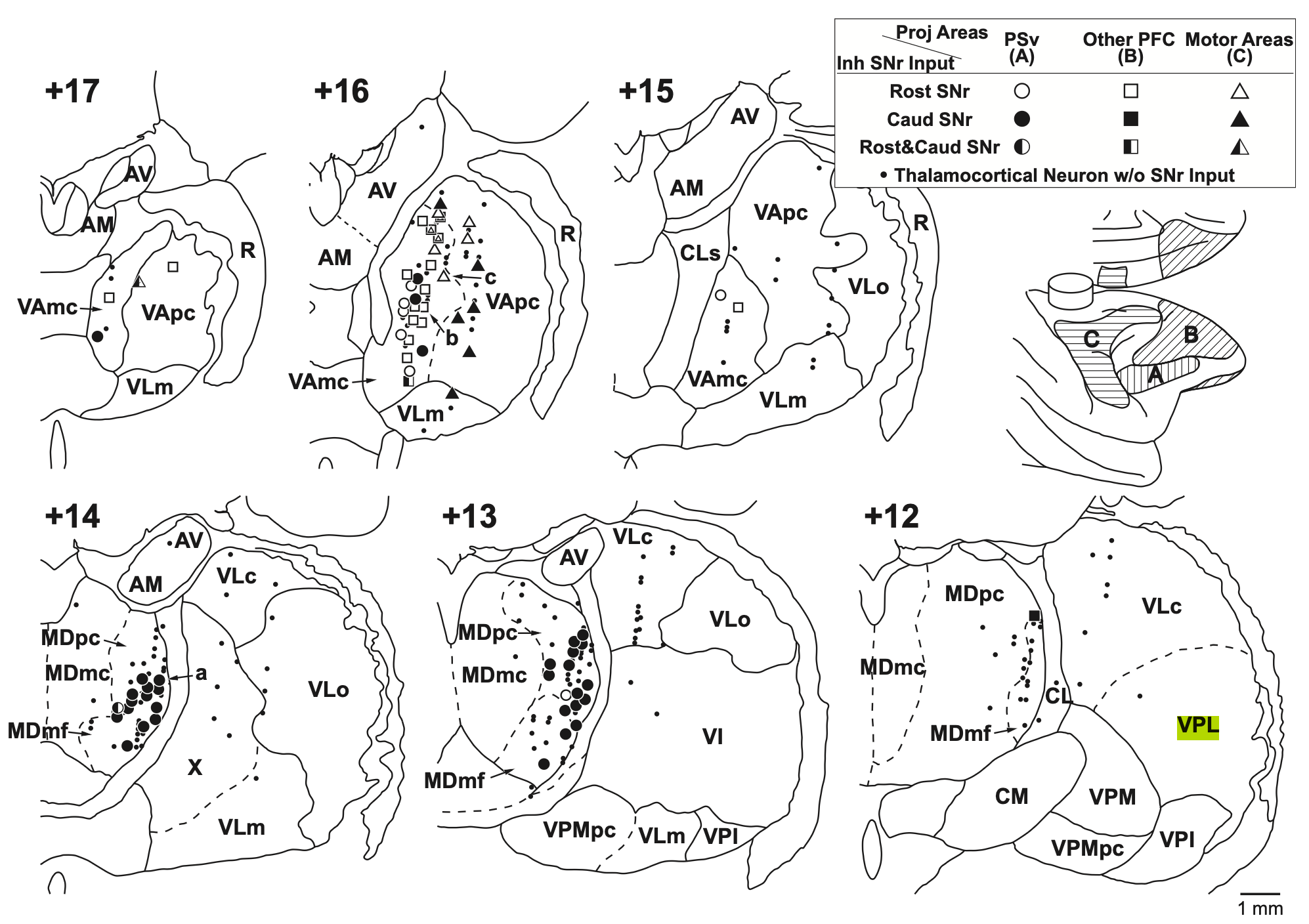
[[#figure_frontal-areas]] ([[@TanibuchiKitanoJinnai09]] より) MD のさまざまな下位区分 (齧歯類の MDm = MDmc、MDc = MDpc、および MDl = MDmf、MDdc)、VLcn (VLm および VLo を含む、ニューロン受信ドライバーとして機能的に定義される) を含む、マカクザルの前頭視床領域の解剖学的組織小脳核からの入力)、およびPFCに投射する霊長類特有のVAmc(VAの内側の大細胞部分)、および齧歯類のVAgpi、我々の用語ではVAsnr(運動野のみに投射し、この霊長類の用語ではVApcおよびLVoの一部にマッピングされます)。
{id=”figure_phillips-cons” style=”高さ:40em”}
![マカクにおける MD 視床と前頭前野 (PFC) の接続パターンの類似性 (これは通常、直接的な相互接続の強さも反映します)。これは、運動前野、dlPFC および vlPFC、および目標駆動領域を明確に描写します。赤とオレンジの領域は、腹側と内側の PFC 目標駆動領域 (つまり、[[Rubicon]] モデルの中核) であり、OFC (13、14、11; 特定の結果)、IL (25; 抽象的な結果)、および PL (32; 統合された報酬コストユーティリティ: 特に 24、25、および 9 とのより広範な相互接続に注意してください) を含みます。黄色 (10) は極前頭皮質で、霊長類に特有で、エピソード記憶と処理の外側ループに重要です。緑色 (47,12) は、側頭葉 (言語、物体など) を制御する vlPFC (腹外側) です。シアン (9、46d、45a) は、行動の空間的および時間的組織化 (広範な計画) のために頭頂葉を制御する dlPFC (背外側) です。青(8)はFEF(前頭眼野)で、PM(運動前野)、SMA(補足運動野)、一次運動野(M1)と同様の接続性を持っています。領域24は、行動の価値(主にコスト)を表すACC(前帯状皮質)であり、運動野だけでなく、目標領域、特に32(PL)とも非常に幅広い相互接続性を持っています。 Phillips et al.、2019 より翻案。](https://raw.githubusercontent.com/compcogneuro/web/main/content/media/fig_thalamus_pfc_simat_phillips_etal_19.png)
[[#figure_phillips-cons]] は、MD 視床接続性 ([[@PhillipsFishKambiEtAl19]]) のパターンに応じたさまざまな PFC 領域の類似構造を示します。これにより、v/mPFC (OFC、IL、PL) の目標駆動野の主要な機能グループ、運動とこれらの目標駆動野を橋渡しして行動価値 (コスト) を表す ACC、後皮質領域 (それぞれ頭頂葉と側頭葉; [[@OReilly10]]) とガイド [[motor]] 前頭葉と相互接続する dlPFC と vlPFC が特定されます。動作(運動前野を示す)。
{id=”figure_frontal-sum” style=”高さ:30em”}
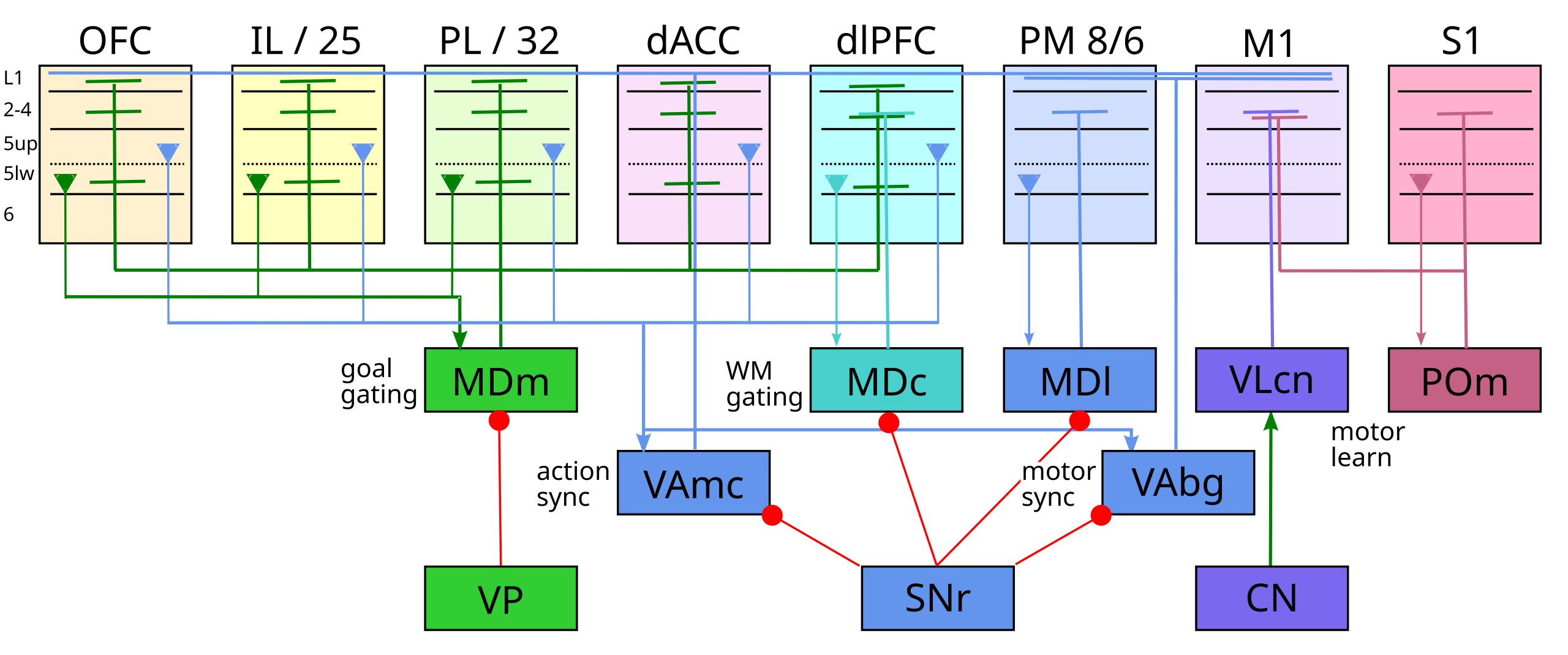
{id=”table_frontal-thal” title=”視床前部領域”} |エリア |機能 |ドライバー |変調器 | BG |ストロングアウト |弱点 |ラミナ |その他のイン |その他のアウト | |———–|————–|—————|———————|——|—–|———–|———-|———————-|————| | 学習| | | | | | | | | | | PULm / POm | S1->M1 学ぶ | S1、脊椎 | M1、M2 | – | M1、M2、SMA.. | |ミッド |脊髄体性感覚 | | | VLcn | CN->M1,M2 学ぶ | CN | M1、M2 | – | M1、M2 | |ミッド | CN | | |医学博士 |予測学習 | (下記参照) | | – | | | | | | | ゲート | (焦点) | | | | | | | | | |午前 | MTL <-> 目標 | ACC、OFC、イリノイ州、PL | d/vlPFC、IOFC | mGPi | 10、11、12、46 | PL、ACC |ミッド | CA3、aSub、BLA、mGPi | vSub、RSC | | MDm(エムシー) |ゴールエンゲージ | OFC、イリノイ州、PL | vlPFC、ACC |副社長 | OFC |イリノイ州PL | S/D、ミッド | IT極、EC、PRh、Sub、BLA | | | MDc (パソコン) |ダウンロード/プラン WM | 10、dlPFC | ACC、PL、PM、SMA | SNr | dlPFC (WM) |首相、SMA |ミッド |頭頂部 |頭頂部 | | MD1 (MF,DC) |アイ/アクトモーター| 8 (FEF)、PM、SMA | | SNr | 8、午後、SMA | |ミッド | SC、頭頂部の集中 | | | ゲート | (拡散) | | | | | | | | | | VAgpi、VAsnr | BG→モーター | | | GPi、SNr | M1、M2、SMA、DM| | | | | | VAmc | BG->dl/モーター | DL、PL、ACC | | SNr | dlPFC、M2、M1 | |ミッド+S/D|頭頂部、一時極 |頭頂部 |
[[#figure_frontal-sum]] および [[#table_frontal-thal]] は、利用可能なすべてのデータ ([[@PhillipsKambiRedinbaughEtAl21]]; [[@TimbieBarbas15]]; [[@XiaoZikopoulosBarbas09]]) に基づいて、上記の 3 つの主要な機能に従って整理された、霊長類およびげっ歯類のすべての運動視床野の全体的な概要を提供します。肺様予測学習とBG駆動のゲーティング/更新のためのMD野の二重機能に注目してください。これは、MDへの投影の80%が第6層のCTニューロンから生じ、20%が深層5b(PTバーストニューロン)から生じることと一致しています([[@XiaoZikopoulosBarbas09]])。
霊長類における MDc の機能は、齧歯動物の場合 (主に OFC を標的とする) と比較して拡張されており、[[PBWM]] と一致して、背側および腹側外側領域 (dlPFC、vlPFC) における作業記憶表現の更新に重要な役割を果たしています ([[@FunahashiBruceGoldman-Rakic89]]; [[@FusterAlexander73]]; [[@TakedaFunahashi02]])。 (前頭前野、大脳基底核ワーキングメモリ) モデル。異なるニューロンが MDc の広大な範囲内の異なる領域をターゲットにしている可能性が高いため、齧歯動物に特有のケースはこれらのニューロンのサブセットによって依然としてサポートされている可能性があります。同様に、霊長類のMD1には、強力なFEF 8aドライバー入力によって駆動される、運動表現のサッケードベースの更新のためのより重要な役割が含まれており、これにより、より広範な運動ゲートが駆動される可能性があります。
霊長類の視床におけるもう 1 つの主要な新しい要素は、視床腹側前部の大細胞区画である VAmc です ([[@PhillipsKambiRedinbaughEtAl21]])。この領域には非常に広範囲に重複する入力があり、入力投影の地形的な組織化は事実上なく、PFC 全体から来て、PFC と運動皮質に広く投影されます。この広範な拡散接続と一致して、マトリックス状のレイヤー 1 出力だけでなく、中間層への投影も送信します。
{id=”figure_pfc-md-va” style=”高さ:40em”}
![視床領域への、および視床領域からの投影の異なるパターン MD および AM 対 VA。 MD と AM は、異なる視床ニューロンが異なる優先標的を持ち、中皮質層と表層の両方に、より局所的に投射するのに対し ([[@MukherjeeLamWimmerEtAl21]] も参照)、VA は両方の標的層にも拡散的に投射します。重要なのは、MD および AM に投射するレイヤー 5 (V) ニューロンは 5b の深部半分 (水色) で発生するのに対し、VA に投射するニューロンは表層半分 (緑色) で発生することです。 [[@^EconomoViswanathanTasicEtAl18]] は、これらの表層ニューロンは準備期間中に活動する可能性が高い一方、深部半分も皮質下に (つまり運動目標に向かって) 投射し、実行中に優先的に活動することを示しました。第 6 層 (VI) ニューロンは視床細胞を投射する中層に優先的に投射しますが、MD および VA ではこれらのニューロンは他の領域から皮質ドライバーも受け取ります (図示せず)。 [[@^XiaoZikopoulosBarbas09]] より。](https://raw.githubusercontent.com/compcogneuro/web/main/content/media/fig_thalamus_pfc_lamina_xiao_etal_09.png)
興味深いことに、VAmc への第 6 層の CT 入力は、皮質の中層に投射する PV を発現するニューロン (コアスタイル) をターゲットとするのに対し、第 5 層の入力は、CB を発現するニューロンをターゲットとする第 5b 層の上部 PT ニューロン ([[@XiaoZikopoulosBarbas09]]; [[@EconomoViswanathanTasicEtAl18]]; [[@HattoxNelson07]]; [[#figure_pfc-md-va]]) から生じます。層 1 の表面層といくつかの深層投影 (マトリックス形式)。 [[@^EconomoViswanathanTasicEtAl18]] で説明されているように、このサブタイプの 5b PT ニューロンは一般に運動の準備と計画中により活発になりますが、第 5b 層の下位 PT ニューロンは運動実行中により活発になり、それに応じて脳幹に投影を送信します。これらの下位サブタイプは、MD および AM 視床に投射するニューロン ([[@XiaoZikopoulosBarbas09]]) であり、[[#figure_pfc-md-va]] に示すように、VA (VAmc、VAgpi、および VAsnr) 領域が MD および AM とは機能的に異なるという考えと一致します。
VAmc の幅広い接続性は、VAbg の接続性と同様に、サッカードや大規模な体の動きなどの主要な運動ベースのイベントに応じて前頭葉運動野と PFC 野にわたる処理を「同期」するのに理想的に適していると思われます。一般に、運動動作の開始時には皮質状態に突然の遷移があり、これらの広範囲の拡散した接続は、これらの遷移を促進する信号を効果的にブロードキャストし、システムが準備活動と実際の運動動作の間で遷移できるようにする可能性があります ([[@ChurchlandShenoy24]]; [[@EconomoViswanathanTasicEtAl18]])。比較的安定した介在状態間の急速な遷移を伴う「メタ安定」神経活動状態に関する文献が増加しており、この動態と一致しています ([[@LaCameraFontaniniMazzucato19]]; [[@RecanatesiPereira-ObilinovicMurakamiEtAl22]])。
[[hippocampus]]、海馬台、内側頭頂葉の脾後皮質 (RSC) を含む内側側頭葉を相互接続する AM の役割は、[[#anterior thalamic areas]] のセクションでもう少し詳しく説明するように、エピソード記憶を目標計画に活用できるようにするために重要です ([[@XiaoZikopoulosBarbas09]] も参照)。
前視床領域
[[@AggletonOMara22]] – キー参照。
- AM: 前内側、MTL と PFC のゴールエリアの間の橋です。[[#table_frontal-thal]] を参照してください。
バースト発射
視床中継細胞には、過分極期間後にバースト発火を引き起こすことができる遅い T 型カルシウムチャネルがあり ([[@LlinasJahnsen82]]; [[@DestexheNeubigUlrichEtAl98]])、このバーストが皮質の注意調節 (つまり、一種の「サーチライト」として) に重要であるというフランシス クリックの仮説を引き起こしました ([[@Crick84]])。しかし、これらの T 型カルシウム チャネルは、活性化するために一定期間の過分極を必要とし、その後、リバウンド発火現象としてバーストが発生します。この過分極は通常、覚醒時ではなく睡眠中に存在し、バーストは睡眠時や非活動状態のときにはるかに多く発生します。
したがって、簡単に説明すると、バーストは、睡眠のほとんどの段階を特徴づける強力な同期活動波 (「徐波」) を引き起こす方法として、睡眠または同様の非活動状態の間にのみ存在します。この同期アクティビティは、有意義な計算に不可欠な [[stable activation]] 状態でスパース [[distributed representations]] をアクティブにする機能を妨害することにより、情報処理を大幅に妨げます。
それにもかかわらず、比較的低い割合で視床バーストが起きており、行動している状態では発生し([[@GuidoWeyand95]]、[[@RamcharanGnadtSherman05]]、[[@BoshraHarrisDoughertyEtAl25]])、それらが機能的に関連しているかどうかという疑問が生じている(例、[[@ZeldenrustWadmanEnglitz18]])。仮説の1つは、それらが一種の「覚醒」効果をもたらし、皮質が以前は活動していなかったとき、または新しい情報によって異なる視床ニューロンが活性化されているときに皮質をさらに興奮させる可能性があるということです(例:[[@Sherman01a]]; [[@Sherman01]])。ただし、[[@^Steriade00]] は次のことを観察しました。
「ウェイクアップ」コール仮説が機能していれば、私たちが徐々に深い睡眠段階に入るにつれて、バーストはより効率的に私たちを目覚めさせるはずです。これは確かに不便でしょう。
ここでの反論は、このバーストと集団全体の活動における広範なレベルの同期が、実際には睡眠状態の処理の重要な推進力であり、覚醒行動中のバーストははるかにまばらで、時間的にもニューロン全体にもより分散しているため、これらのレベルの同期発火は引き起こされないということです。
バースト活動を操作し、皮質標的への影響を測定した研究から、バーストが実際に、一連の急速な興奮性入力から予想されるような種類の興奮性の影響を皮質に及ぼすことが明らかである([[@AlittoRathbunVandeleestEtAl19]]; [[@SwadlowGusev01]]; [[@BoshraHarrisDoughertyEtAl25]])。
したがって、T 型カルシウム チャネルの特性と、バースト興奮による大きな下流への影響を総合すると、「新しい」入力によって引き起こされるバーストによってある程度の注意力が増強されることは理にかなっています。ただし、T 型チャネルを活性化するには過分極が必要であるため、何らかの継続的な活動が存在する場合は常に、バーストが主要な原因となるべきではありません。
バースト確率を決定する際のベースライン活動レベルの重要性は、一次感覚視床(一般に高レベルの感覚主導型活動を有する)と、実行されるタスクの種類に応じてよりまばらに活性化される可能性がある高次視床領域を比較した研究によって示されています。初期の報告の 1 つ ([[@GuidoWeyand95]]) では、起きていて行動している猫の一次視覚野におけるバースト確率が 1% 未満であることが示されています。上記の説明と一致して、彼らはまた、これらのほとんどが視覚刺激の開始時に発生したと報告しています。
[[@^RamcharanGnadtSherman05]]は、覚醒しているがほとんど行動していないサル(頭を固定し、何もせずに座っているか、単純な固定を保持している)において、一次感覚視床(LGN、VP、MGNv)よりも高次視床(骨盤およびMD)のニューロンのバーストのレベルが高い(10〜20%)と報告しました。 [[@^BoshraHarrisDoughertyEtAl25]] は、起きて行動しているサルの胸骨の破裂はアイドル期間に発生する可能性が高いが、出席している場所ではわずかに増加することを示しました。彼らは電気刺激によって引き起こされるバーストの効果を強調しましたが、純粋に自然主義的なバーストが明確な行動的影響を持っているかどうかは明らかではありませんでした。
結論として、視床破裂は主に睡眠に関連している可能性が高いため、モデルから安全に省略でき、「省略のエラー」として小さなペナルティが発生する可能性があります。以前は不活性だった視床ニューロンからの新たな活動は、皮質ニューロンの駆動にすでに大きな影響を与えている可能性が高いため、行動への影響は比較的穏やかであると考えられます。