compcogneuro/web: vestibular
このページは外部資料の日本語訳です。原文の見出し順と本文順を保ち、コード・URL・出典表記はできるだけ原形のまま残しています。
出典とライセンス
原典: https://github.com/compcogneuro/web/blob/main/content/vestibular.md
ライセンス: Text: CC BY 4.0; code: BSD 3-Clause。このページは日本語翻訳であり、変更点は翻訳とサイト内整形です。
+++ Categories = [“Neuroscience”] bibfile = “ccnlab.json” +++ 前庭系は、脳による [[space]] および [[motor]] 制御の表現にとって重要な感覚経路であり、耳の内部にある精巧な感覚器官のシステムを介して、頭の加速度、速度、位置の感覚読み取りを提供します。これらの信号は、[[cerebellum]] の体性感覚信号、固有受容信号、および視覚信号と統合され、モーター制御に不可欠な現在の頭の位置を正確かつ効果的に表現します。運動動作は主に、頭部にある感覚系 (目、耳、鼻) から生じる感覚入力によって駆動されるため、適切に調整された運動動作を生成するには、現在の頭の向きと運動パラメータを知ることが不可欠です。
さらに、広い空間を移動する私たちの能力は、「頭の方向」表現として知られる、より大きな空間環境内での体の向きの継続的に更新される表現に決定的に依存しています。これらは、前庭核の深部から発生する入力から計算されます。
{id=”figure_system” style=”高さ:35em”}
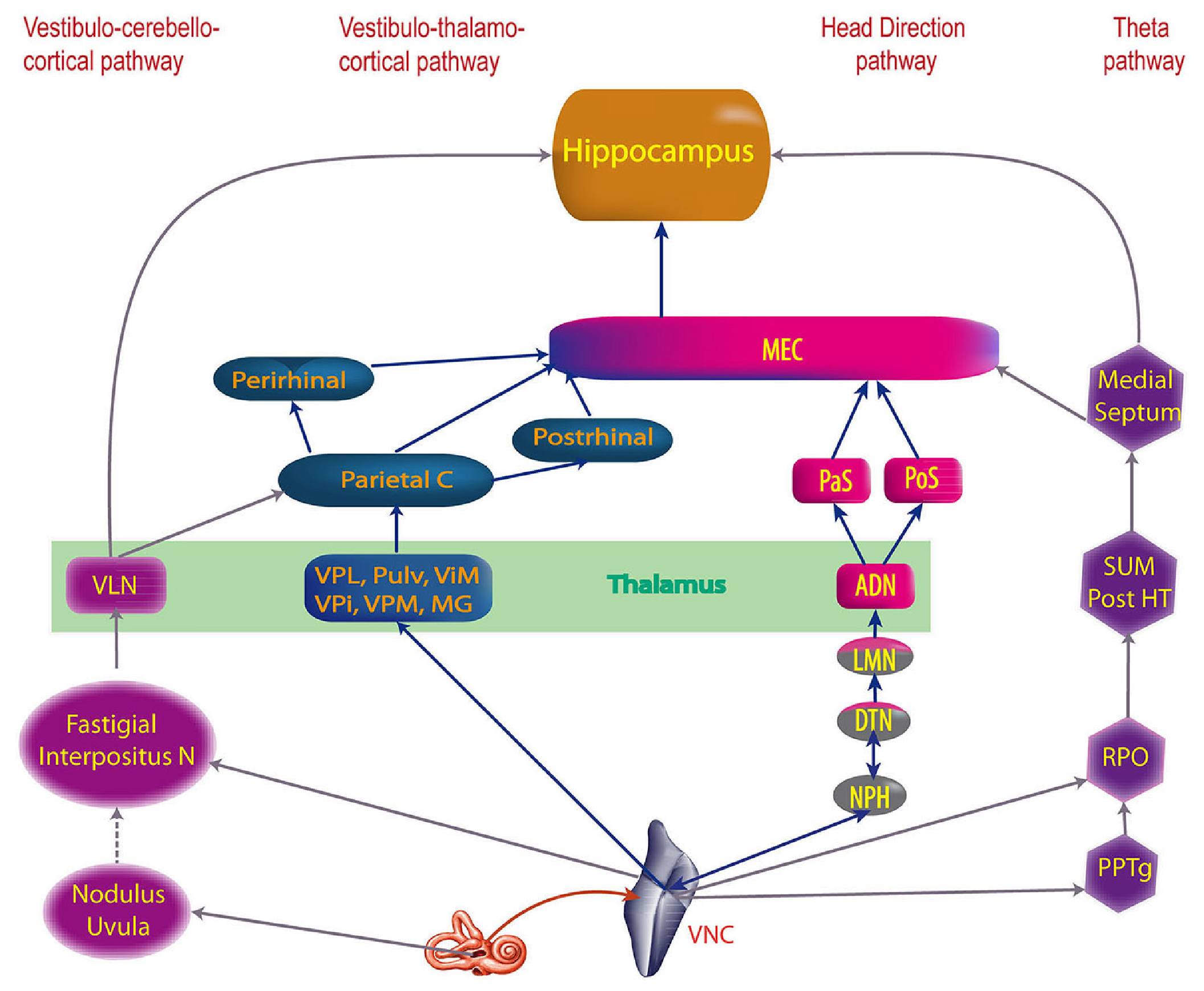
[[#figure_system]] ([[@HitierBesnardSmith14]] より) は、前庭系とその脳の残りの部分への投影の概要を示し、4 つの主要な管に編成されています。
-
[[Cerebellum]]: 小脳の最も古い部分 [[evolution evolutionarily]] は、fastigial および interpositus (介在) 深小脳核 (DCN) を介して、前庭系と強く双方向に相互接続されています。運動処理における前庭信号の中心的な役割は、これらのシステムの大規模な相互接続に反映されています。 -
[[neocortex]] への直接: 新皮質、特に頭頂葉の体性感覚系、[[space spatial]] および [[motor]] 系への最も直接的な投射は、後腹側 (VP) および内側膝状突起 (MG、内側) の領域を含む一連の腹側 [[thalamus thalamic]] 核を経由します。部分)核。 VP は一次体性感覚視床、MG は一次聴覚視床であり、どちらも感覚情報を皮質に伝える強力な皮質下ドライバー入力を持っています。賢明なことに、前庭系はこれらの両方に寄与しています。 -
頭の方向: 前庭 (および運動) ソースからの頭方向信号の計算に関与するいくつかの中脳核があり、(グッデンの) 背側被蓋核 (DTN)_ は、頭と体の動きの信号のさまざまな表現を持つ収束領域であり、[[space#head direction cells]] が最初に出現する外側乳頭核 (LMN)_ に統合されます。次に、LMN は、AD (前背側) 視床に強力なドライバー入力を提供し、視床はこれらの信号を、より広範な [[hippocampus hippocampal]] システムの一部である臍帯領域に送信します。これらの頭方向細胞と海馬台のより統合的な空間表現は、海馬の記憶機能に中核となる空間アンカーを提供します。 - [[Theta rhythm]]: 前庭系は、乳頭上 (SUM) および内側中隔シータリズム ネットワークにも入力を提供します。これらも (グッデンの) 腹側被蓋核 (VTN) からのリズム活動によって駆動されます。
前庭伝達
{id=”figure_transduction” style=”高さ:25em”}
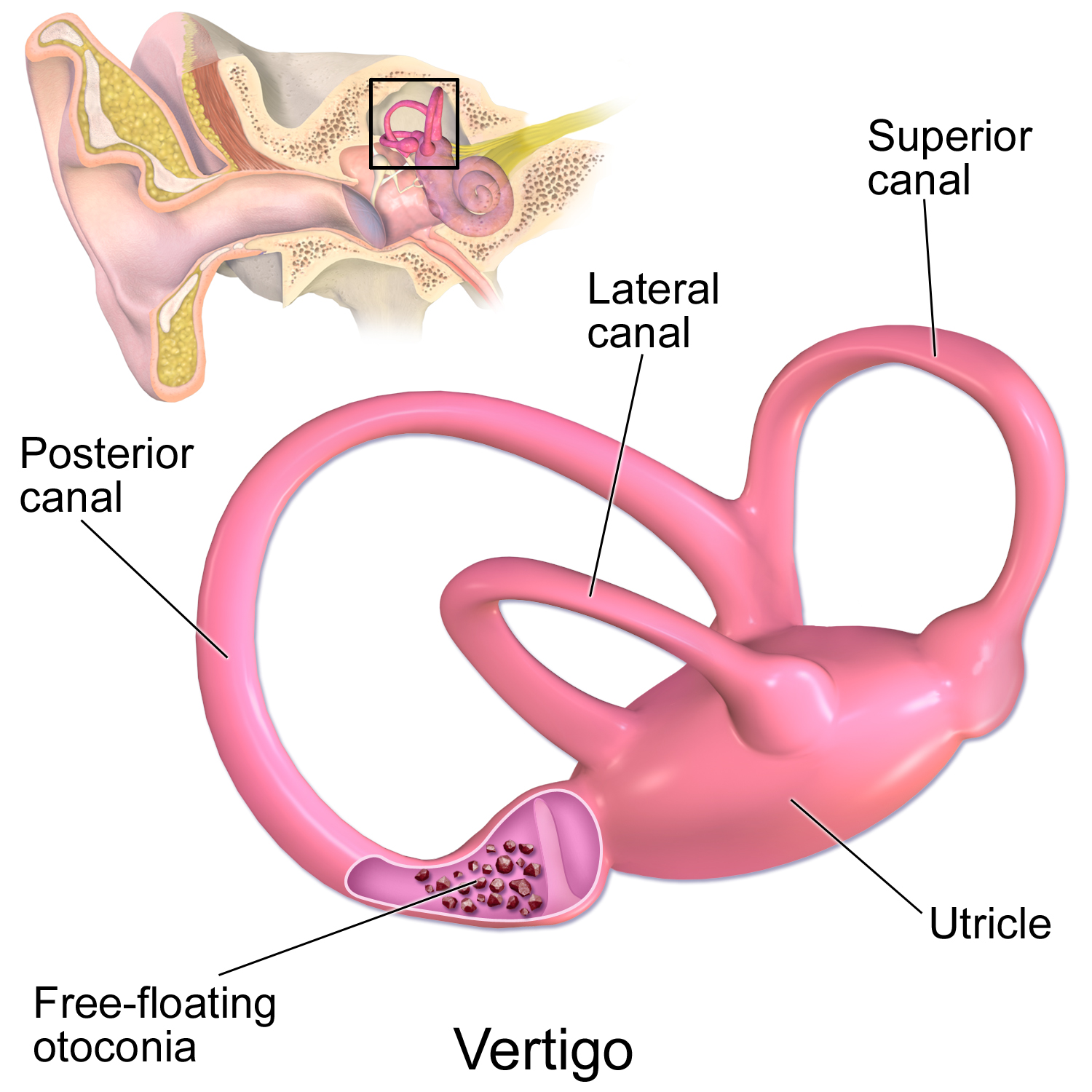
頭の動きの神経発火信号への変換は、前庭器官 ([[#figure_transduction]]) を介して行われます。ここで、三半規管は 3 つの異なる軸の回転運動に敏感ですが、耳石器官は直線加速度 (水平方向の 球形、垂直方向の 球形) に敏感です。
前庭核 (VNC)
VNC の前庭核は、傍孤立核 (Psol) に加えて、下行核、側方核、内側核、上庭核 (DVN、LVN、MVN、SVN) に細分できます。これらの違いは、一次前庭信号 ([[@Barmack23]]) からの求心性入力よりも、その投影ターゲットの方が大きく異なります。これらの一次求心性入力はもっぱら同側性(例、左耳から左半球へ)ですが、核ニューロンからの出力経路は同側性または対側性に投射することができます。
異なる核間には大規模な半球横断接続があり、一般にこれらのニューロン間に [[opponent]] 関係が確立されます。一方の興奮は他方の比例した抑制につながります。これは、水平方向の信号を処理するニューロン (三半規管と卵形嚢耳石) に特有です。
これらの VNC ニューロンは、一次前庭入力に加えて、副光学系 (AOS) からの視覚運動信号と首の固有受容信号を受け取ります。
VNC 内のさまざまな亜核は、機能的に強く組織化されておらず、複数の異なる亜核内のニューロンが、小脳虫や傍綿状筋、目の制御のための外眼筋 (例: [[cerebellum#vestibulo-ocular reflex]])、MDJ 内のさまざまな領域 (例: カハール間質核、傍筋膜下複合体、核など) を含むさまざまな標的領域に投影されています。 Darkschewitsch の)、および [[thalamus]] (VP)。